- Microtubule
-
 Space-filling model of a microtubule segment derived from cryo-electron microscopy. The protofilaments are seen running along the axis of the segment. The microtubule (+) end is towards the top of the image.[1]
Space-filling model of a microtubule segment derived from cryo-electron microscopy. The protofilaments are seen running along the axis of the segment. The microtubule (+) end is towards the top of the image.[1]
Microtubules are a component of the cytoskeleton. These rope-like polymers of tubulin can grow as long as 25 micrometers and are highly dynamic. The outer diameter of microtubule is about 25 nm. Microtubules are important for maintaining cell structure, providing platforms for intracellular transport, forming the spindle during mitosis, as well as other cellular processes.[2] There are many proteins that bind to the microtubule, including motor proteins such as kinesin and dynein, severing proteins like katanin, and other proteins important for regulating microtubule dynamics. [3] [4]
Contents
Structure
Microtubules are polymers of α- and β-tubulin dimers. The tubulin dimers polymerize end to end in protofilaments. Typically 13 protofilaments associate laterally to form the microtubule, although microtubules containing 14 protofilaments have been seen in vitro.The protofilaments arrange themselves in an imperfect helix with one turn of the helix containing 13 tubulin dimers each from a different protofilament. The image above illustrates a small section of microtubule, a few αβ dimers in length.
Another important feature of microtubule structure is polarity. Tubulin polymerizes end to end with the α subunit of one tubulin dimer contacting the β subunit of the next. Therefore, in a protofilament, one end will have the α subunit exposed while the other end will have the β subunit exposed. These ends are designated the (−) and (+) ends, respectively. The protofilaments bundle parallel to one another, so in a microtubule, there is one end, the (+) end, with only β subunits exposed while the other end, the (−) end, only has α subunits exposed. The (-) end is capped so elongation of the microtubule occurs from the (+) direction.
Organization within cells
Microtubules are nucleated and organized by the microtubule organizing centers (MTOCs), such as centrioles and basal bodies. Microtubules are part of a structural network (the cytoskeleton) within the cell's cytoplasm, but, in addition to structural support, microtubules take part in many other processes. They are capable of growing and shrinking in order to generate force, and there are also motor proteins that allow organelles and other cellular factors to move along the microtubule. A notable structure involving microtubules is the mitotic spindle used by eukaryotic cells to segregate their chromosomes correctly during cell division. Microtubules are also part of the cilia and flagella of eukaryotic cells (prokaryote flagella are entirely different).
The process of mitosis is facilitated by a subgroup of microtubules known as astral microtubules, defined as a microtubule originating from the centrosome that does not connect to a kinetochore. Astral microtubules develop in the actin skeleton and interact with the cell cortex to aid in spindle orientation. They are organized into radial arrays around the centrosomes. The turn-over rate of this population of microtubules is higher than any other population. Astral microtubules function in concert with specialized dynein motors, which are oriented with the light chain portion attached to the cell membrane and the dynamic portion attached to the microtubule. This allows for dynein contraction to pull the centrosome toward the cell membrane, thus assisting in cytokinesis. Astral microtubules are not required for the progression of mitosis, but they are required to ensure the fidelity of the process; they are required for the correct positioning and orientation of the mitotic spindle apparatus. They are also involved in determination of cell division site based on the geometry and polarity of the cells. The maintenance of astral microtubules is dependent on the integrity of centrosome. It is also dependent on several microtubule-associated proteins such as EB1 and Adenomatous Polyposis Coli (APC).
Nucleation and growth
Polymerization of microtubules is nucleated in a microtubule organizing center. Contained within the MTOC is another type of tubulin, γ-tubulin, which is distinct from the α and β subunits, which compose the microtubules themselves. The γ-tubulin combines with several other associated proteins to form a circular structure known as the "γ-tubulin ring complex." This complex acts as a scaffold for α/β tubulin dimers to begin polymerization; it acts as a cap of the (−) end while microtubule growth continues away from the MTOC in the (+) direction.
Dynamic instability
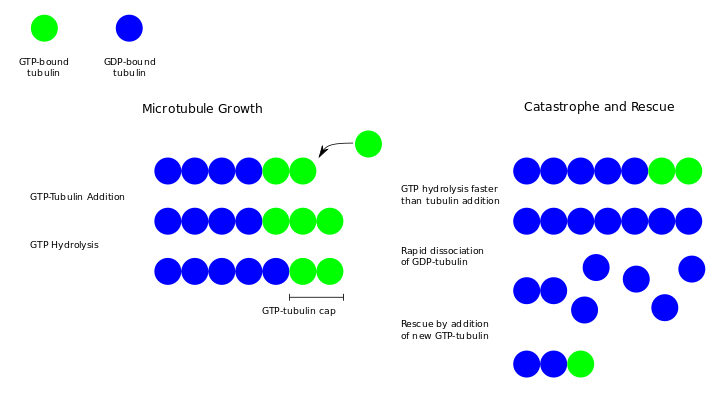
Dynamic instability refers to the coexistence of assembly and disassembly at the (+) end of a microtubule. The microtubule can switch between the growing and shrinking phases dynamically at this region.[5] During polymerization, both the α- and β-subunits of the tubulin dimer are bound to a molecule of GTP. While the GTP bound to α-tubulin is stable, the GTP bound to β-tubulin may be hydrolyzed to GDP shortly after assembly. The kinetics of GDP-tubulin are different from those of GTP-tubulin; GDP-tubulin is prone to depolymerization. A GDP-bound tubulin subunit at the tip of a microtubule will fall off, though a GDP-bound tubulin in the middle of a microtubule cannot spontaneously pop out. Since tubulin adds onto the end of the microtubule only in the GTP-bound state, there is a cap of GTP-bound tubulin at the tip of the microtubule, protecting it from disassembly. When hydrolysis catches up to the tip of the microtubule, it begins a rapid depolymerization and shrinkage. This switch from growth to shrinking is called a catastrophe. GTP-bound tubulin can begin adding to the tip of the microtubule again, providing a new cap and protecting the microtubule from shrinking. This is referred to as "rescue."[6]
In vivo microtubule dynamics vary considerably. Assembly, disassembly, and catastrophe rates depend on which microtubule-associated proteins (MAPs) are present.
Chemical effects on microtubule dynamics
Microtubule dynamics can also be altered by drugs.
- For example, the taxane drug class (e.g., paclitaxel or docetaxel), used in the treatment of cancer, blocks dynamic instability by stabilizing GDP-bound tubulin in the microtubule. Thus, even when hydrolysis of GTP reaches the tip of the microtubule, there is no depolymerization and the microtubule does not shrink back.
- Nocodazole, vincristine, and Colchicine have the opposite effect, blocking the polymerization of tubulin into microtubules.
- Eribulin binds to the (+) growing end of the microtubules.
Motor proteins
In addition to movement generated by the dynamic instability of the microtubule itself, the fibers are substrates along which motor proteins can move. The major microtubule motor proteins are kinesin, which moves toward the (+) end of the microtubule, and dynein, which moves toward the (−) end.
Postulated role in consciousness
In their controversial Orch-OR theory of consciousness, Roger Penrose and Stuart Hameroff postulate that microtubules in neurons conduct quantum-level manipulations of matter, which produces consciousness, based partially on some observations of Gamma Synchrony that indicate that information may propagate through the brain much faster than a chemically mediated neural network would physically permit. Max Tegmark disputes the relevance of these observations, and the matter remains open to debate. David Chalmers[7] argues that quantum theories of consciousness suffer from the same weakness as more conventional theories. Just as he argues that there is no particular reason why particular macroscopic physical features in the brain should give rise to consciousness, he also thinks that there is no particular reason why a particular quantum feature, such as the EM field in the brain, should give rise to consciousness either. While at least one researcher claims otherwise, Jeffrey Gray states in his book Consciousness: Creeping up on the Hard Problem, that tests looking for the influence of electromagnetic fields on brain function have been universally negative in their result.[8]
Additional images
-

Proteins in different cellular compartments and structures tagged with green fluorescent protein.
References
- ^ Li H, DeRosier DJ, Nicholson WV, Nogales E, Downing KH (October 2002). "Microtubule structure at 8 Å resolution". Structure 10 (10): 1317–28. doi:10.1016/S0969-2126(02)00827-4. PMID 12377118.
- ^ Desai A.; and Mitchison TJ; (1997). "Microtubule polymerization dynamics.". Annu Rev Cell Dev Biol 13: 83–117. doi:10.1146/annurev.cellbio.13.1.83. PMID 9442869.
- ^ Vale RD (Feb 2003). "The molecular motor toolbox for intracellular transport.". Cell 112 (4): 467–80. doi:10.1016/S0092-8674(03)00111-9. PMID 12600311.
- ^ Howard J; Hyman AA; (Feb 2007). "Microtubule polymerases and depolymerases.". Curr Opin Cell Biol 19 (1): 31–5. doi:d10.1016/j.ceb.2006.12.009.
- ^ Karp, Gerald (2005). Cell and Molecular Biology: Concepts and Experiments. USA: John Wiley & Sons. p. 355. ISBN 0-471-46580-1.
- ^ Mitchison T, Kirschner M (15-21 November 1984). "Dynamic instability of microtubule growth". Nature 312 (5991): 237–42. doi:10.1038/312237a0. PMID 6504138.
- ^ David Chalmers. The Conscious Mind: In Search of a Fundamental Theory. ISBN 0195105532.
- ^ Jeffrey Gray (2004). Consciousness: Creeping up on the Hard Problem. Oxford University Press. ISBN 0-19-852090-5.
Proteins of the cytoskeleton Human I (MYO1A, MYO1B, MYO1C, MYO1D, MYO1E, MYO1F, MYO1G, MYO1H) · II (MYH1, MYH2, MYH3, MYH4, MYH6, MYH7, MYH7B, MYH8, MYH9, MYH10, MYH11, MYH13, MYH14, MYH15, MYH16) · III (MYO3A, MYO3B) · V (MYO5A, MYO5B, MYO5C) · VI (MYO6) · VII (MYO7A, MYO7B) · IX (MYO9A, MYO9B) · X (MYO10) · XV (MYO15A) · XVIII (MYO18A, MYO18B) · LC (MYL1, MYL2, MYL3, MYL4, MYL5, MYL6, MYL6B, MYL7, MYL9, MYLIP, MYLK, MYLK2, MYLL1)OtherOtherEpithelial keratins
(soft alpha-keratins)Hair keratins
(hard alpha-keratins)Ungrouped alphaNot alphaType 3Type 4Type 5Microtubules/
MAPsOtherOtherNonhuman see also cytoskeletal defects
B strc: edmb (perx), skel (ctrs), epit, cili, mito, nucl (chro)Categories:- Cell anatomy
- Cytoskeleton



Wikimedia Foundation. 2010.