- Myosin
-
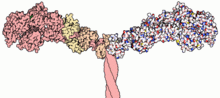 Part of the myosin II structure. Atoms in the heavy chain are colored red on the left-hand side, and atoms in the light chains are colored orange and yellow.
Part of the myosin II structure. Atoms in the heavy chain are colored red on the left-hand side, and atoms in the light chains are colored orange and yellow.
Myosins comprise a family of ATP-dependent motor proteins and are best known for their role in muscle contraction and their involvement in a wide range of other eukaryotic motility processes. They are responsible for actin-based motility. The term was originally used to describe a group of similar ATPases found in striated and smooth muscle cells.[1] Following the discovery by Pollard and Korn of enzymes with myosin-like function in Acanthamoeba castellanii, a large number of divergent myosin genes have been discovered throughout eukaryotes. Thus, although myosin was originally thought to be restricted to muscle cells (hence, "myo"), there is no single "myosin" but rather a huge superfamily of genes whose protein products share the basic properties of actin binding, ATP hydrolysis (ATPase enzyme activity), and force transduction. Virtually all eukaryotic cells contain myosin isoforms. Some isoforms have specialized functions in certain cell types (such as muscle), while other isoforms are ubiquitous. The structure and function of myosin is strongly conserved across species, to the extent that rabbit muscle myosin II will bind to actin from an amoeba.[2]
Structure and function
Domains
Most myosin molecules are composed of a head, neck, and tail domain.
- The head domain binds the filamentous actin, and uses ATP hydrolysis to generate force and to "walk" along the filament towards the barbed (+) end (with the exception of myosin VI, which moves towards the pointed (-) end).
- the neck domain acts as a linker and as a lever arm for transducing force generated by the catalytic motor domain. The neck domain can also serve as a binding site for myosin light chains which are distinct proteins that form part of a macromolecular complex and generally have regulatory functions.
- The tail domain generally mediates interaction with cargo molecules and/or other myosin subunits. In some cases, the tail domain may play a role in regulating motor activity.
Power stroke
Main article: Muscle contractionMultiple myosin II molecules generate force in skeletal muscle through a power stroke mechanism fuelled by the energy released from ATP hydrolysis.[3] The power stroke occurs at the release of phosphate from the myosin molecule after the ATP hydrolysis while myosin is tightly bound to actin. The effect of this release is a conformational change in the molecule that pulls against the actin. The release of the ADP molecule and binding of a new ATP molecule will release myosin from actin. ATP hydrolysis within the myosin will cause it to bind to actin again to repeat the cycle. The combined effect of the myriad power strokes causes the muscle to contract.
Nomenclature, evolution, and the family tree
The wide variety of myosin genes found throughout the eukaryotic phyla were named according to different schemes as they were discovered. The nomenclature can therefore be somewhat confusing when attempting to compare the functions of myosin proteins within and between organisms.
Skeletal muscle myosin, the most conspicuous of the myosin superfamily due to its abundance in muscle fibers, was the first to be discovered. This protein makes up part of the sarcomere and forms macromolecular filaments composed of multiple myosin subunits. Similar filament-forming myosin proteins were found in cardiac muscle, smooth muscle, and non-muscle cells. However, beginning in the 1970s researchers began to discover new myosin genes in simple eukaryotes[1] encoding proteins that acted as monomers and were therefore entitled Class I myosins. These new myosins were collectively termed "unconventional myosins"[4] and have been found in many tissues other than muscle. These new superfamily members have been grouped according to phylogenetic relationships derived from a comparison of the amino acid sequences of their head domains, with each class being assigned a Roman numeral[5][6][7][8](see phylogenetic tree). The unconventional myosins also have divergent tail domains, suggesting unique functions.[9] The now diverse array of myosins likely evolved from an ancestral precursor (see picture).
 Myosin unrooted phylogenetic tree
Myosin unrooted phylogenetic treeAnalysis of the amino acid sequences of different myosins shows great variability among the tail domains but strong conservation of head domain sequences. Presumably this is so the myosins may interact, via their tails, with a large number of different cargoes, while the goal in each case - to move along actin filaments - remains the same and therefore requires the same machinery in the motor. For example, the human genome contains over 40 different myosin genes.
These differences in shape also determine the speed at which myosins can move along actin filaments. The hydrolysis of ATP and the subsequent release of the phosphate group causes the "power stroke," in which the "lever arm" or "neck" region of the heavy chain is dragged forward. Since the power stroke always moves the lever arm by the same angle, the length of the lever arm determines the displacement of the cargo relative to the actin filament. A longer lever arm will cause the cargo to traverse a greater distance even though the lever arm undergoes the same angular displacement - just as a person with longer legs can move farther with each individual step. The velocity of a myosin motor depends upon the rate at which it passes through a complete kinetic cycle of ATP binding to the release of ADP.
Myosin classes
Myosin I
Myosin I functions as monomer and functions in vesicle transport.[10] It has step size of 10 nm and has been implicated as being responsible for the adaptation response of the stereocilia in the inner ear.[11]
Myosin II
 Sliding filament model of muscle contraction.
Sliding filament model of muscle contraction.Myosin II (also known as conventional myosin) is the myosin type responsible for producing muscle contraction in muscle cells.
- Myosin II contains two heavy chains, each about 2000 amino acids in length, which constitute the head and tail domains. Each of these heavy chains contains the N-terminal head domain, while the C-terminal tails take on a coiled-coil morphology, holding the two heavy chains together (imagine two snakes wrapped around each other, such as in a caduceus). Thus, myosin II has two heads. The intermediate neck domain is the region creating the angle between the head and tail.[12] In smooth muscle, there is a single gene (MYH11[13]) that codes for the heavy chains myosin II, but there are splice variants of this gene that result in four distinct isoforms.[12]
- It also contains 4 light chains, resulting in 2 per head, weighing 20 (MLC20) and 17 (MLC17) kDa.[12] These bind the heavy chains in the "neck" region between the head and tail.
- The MLC20 is also known as the regulatory light chain and actively participates in muscle contraction.[12]
- The MLC17 is also known as the essential light chain.[12] Its exact function is unclear, but it's believed that it contributes to the structural stability of the myosin head along with MLC20.[12] Two variants of MLC17 (MLC17a/b) exist as a result of alternate splicing at the MLC17 gene.[12]
In muscle cells, the long coiled-coil tails of the individual myosin molecules join together, forming the thick filaments of the sarcomere. The force-producing head domains stick out from the side of the thick filament, ready to walk along the adjacent actin-based thin filaments in response to the proper chemical signals.
Further information: Muscle contractionMyosin III
Myosin III is a poorly understood member of the myosin family. It has been studied in vivo in the eyes of Drosophila, where it is thought to play a role in phototransduction.[14] A human homologue gene for myosin III, MYO3A, has been uncovered through the Human Genome Project and is expressed in the retina and cochlea.[15]
Myosin IV
Myosin IV has a single IQ motif and a tail that lacks any coiled-coil forming sequence. It has homology similar to the tail domains of Myosin VII and XV.[16]
Myosin V
Myosin V is an unconventional myosin motor, which is functional as a dimer. It translocates (walks) along actin filaments traveling towards the barbed end (+ end) of the filaments. Myosin V was thought to be critical in vesicle movement from the center of the cell to the periphery, but has been shown to be more like a dynamic tether retaining vesicles and organelles in the actin-rich periphery of cells.
Myosin VI
 Pre-powerstroke state of myosin VI from PDB 2V26. [17]
Pre-powerstroke state of myosin VI from PDB 2V26. [17]Myosin VI is an unconventional myosin motor, which is primarily functional as a dimer but also acts as a nonprocessive monomer. It walks along actin filaments travelling towards the minus end of the filaments.[18] Myosin VI is thought to transport endocytic vesicles into the cell.[19]
Myosin VII
Myosin VII is an unconventional myosin with two FERM domains in the tail region. It has a very short coiled-coiled region. Myosin VII is required for phagocytosis in Dictyostelium discoideum, spermatogenesis in C. elegans and stereocilia formation in mice and zebrafish.[20]
Myosin VIII
Myosin VIII is a plant-specific myosin linked to cell division;[21] specifically, it is involved in regulating the flow of cytoplasm between cells[22] and in the localisation of vesicles to the phragmoplast.[23]
Myosin IX
Myosin IX is a single-headed, minus-end directed group of motor proteins. The movement mechanism for this myosin is poorly understood.[24]
Myosin X
Myosin X is an unconventional myosin motor, which is functional as a dimer. In mammalian cells the motor is found to localize to filopodia. Myosin X walks towards the barbed ends of filaments. Some research suggests that it preferentially walks on bundles of actin, rather than single filaments. It is the first myosin motor to exhibit this behavior.
Myosin XI
Myosin XI directs the movement of organelles as plastids and mitochondria in plant cells.[25] It is responsible of the light-directed movement of chloroplasts according to light intensity and the formation of stromules interconnecting different plastids.
Myosin XII
Myosin XIII
Myosin XIV
This myosin group has been found in the Apicomplexa phylum.[26] The myosins localise to plasma membranes of the intracellular parasites and may then be involved in the cell invasion process.[27]
This myosin is also found in the ciliated protozoan Tetrahymena thermaphila. Known functions include: transporting phagosomes to the nucleus and perturbing the developmentally regulated elimination of the macronucleus during conjugation.
Myosin XV
Myosin XV is a motor that is involved in the structuring and motion of stereocilia of in the inner ear. It is thought to be functional as a monomer.
Myosin XVI
Myosin XVII
Myosin XVIII
Genes in humans
Note that not all of these genes are active.
- Class I: MYO1A, MYO1B, MYO1C, MYO1D, MYO1E, MYO1F, MYO1G, MYO1H
- Class II: MYH1, MYH2, MYH3, MYH4, MYH6, MYH7, MYH7B, MYH8, MYH9, MYH10, MYH11, MYH13, MYH14, MYH15, MYH16
- Class III: MYO3A, MYO3B
- Class V: MYO5A, MYO5B, MYO5C
- Class VI: MYO6
- Class VII: MYO7A, MYO7B
- Class IX: MYO9A, MYO9B
- Class X: MYO10
- Class XV: MYO15A
- Class XVIII: MYO18A, MYO18B
Myosin light chains are distinct and have their own properties. They are not considered "myosins" but are components of the macromolecular complexes that make up the functional myosin enzymes.
Paramyosin
Paramyosin is a large 93-115kDa muscle protein that has been described in a number of diverse invertebrate phyla.[28] It is thought that invertebrate thick filaments are composted of an inner paramyosin core that is surrounded by myosin. The myosin interacts with actin resulting in fibre contraction.[29] Paramyosin is found in many different invertebrate species, for example, Brachiopoda, Sipunculidea, Nematoda, Annelida, Mollusca, Arachnida, Insecta.[28] Paramyosin is responsible for the "catch" mechanism that enables sustained contraction of muscles with very little energy expenditure, such that a clam can remain closed for extended periods.
See also
References
- ^ a b Pollard, Thomas D.; Korn, Edward D. (1973). "Acanthamoeba myosin. I. Isolation from Acanthamoeba castellanii of an enzyme similar to muscle myosin.". The Journal of biological chemistry 248 (13): 4682–90. PMID 4268863. http://www.jbc.org/content/248/13/4682.abstract.
- ^ McMahon, T. A. 1984. Muscles, Reflexes and Locomotion. 1st Edition. Princeton University Press. ISBN 978-0691023762
- ^ Tyska, Matthew J.; Warshaw, David M. (2002). "The myosin power stroke". Cell Motility and the Cytoskeleton 51 (1): 1–15. doi:10.1002/cm.10014. PMID 11810692.
- ^ Cheney, Richard E.; Mooseker, Mark S. (1992). "Unconventional myosins". Current opinion in cell biology 4 (1): 27–35. doi:10.1016/0955-0674(92)90055-H. PMID 1558751.
- ^ Cheney, Richard E.; Riley, Margaret A.; Mooseker, Mark S. (1993). "Phylogenetic analysis of the myosin superfamily". Cell motility and the cytoskeleton 24 (4): 215–23. doi:10.1002/cm.970240402. PMID 8477454.
- ^ Goodson, HV (1994). "Molecular evolution of the myosin superfamily: application of phylogenetic techniques to cell biological questions". Society of General Physiologists series 49: 141–57. PMID 7939893.
- ^ Hodge, Tony; Cope, M. Jamie T. V. (2000). "A myosin family tree". Journal of cell science 113 Pt 19: 3353–4. PMID 10984423.
- ^ Berg, Jonathan S.; Powell, Bradford C.; Cheney, Richard E. (2001). "A Millennial Myosin Census". Molecular biology of the cell 12 (4): 780–94. PMC 32266. PMID 11294886. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=32266.
- ^ Oliver, T. N.; Berg, J. S.; Cheney, R. E. (1999). "Tails of unconventional myosins". Cellular and molecular life sciences 56 (3–4): 243–57. doi:10.1007/s000180050426. PMID 11212352.
- ^ Sutherland Macive (June 4, 2003). "Myosin I". http://www.bms.ed.ac.uk/research/others/smaciver/Myosin%20I.htm. Retrieved 2007-05-23.
- ^ Batters, Christopher; Arthur, Christopher P; Lin, Abel; Porter, Jessica; Geeves, Michael A; Milligan, Ronald A; Molloy, Justin E; Coluccio, Lynne M (2004). "Myo1c is designed for the adaptation response in the inner ear". The EMBO Journal 23 (7): 1433–40. doi:10.1038/sj.emboj.7600169. PMC 391074. PMID 15014434. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=391074.
- ^ a b c d e f g Aguilar, H. N.; Xiao, S.; Knoll, A. H.; Yuan, X. (2010). "Physiological pathways and molecular mechanisms regulating uterine contractility". Human Reproduction Update 16 (6): 725–744. doi:10.1093/humupd/dmq016. JSTOR 1306737. PMID 20551073.
- ^ Matsuoka, R.; Yoshida, M. C.; Furutani, Y.; Imamura, S. I.; Kanda, N.; Yanagisawa, M.; Masaki, T.; Takao, A. (1993). "Human smooth muscle myosin heavy chain gene mapped to chromosomal region 16q12". American Journal of Medical Genetics 46 (1): 61. doi:10.1002/ajmg.1320460110. ISBN 1320460110. PMID 7684189.
- ^ http://www.bms.ed.ac.uk/research/others/smaciver/Myosin%20III.htm
- ^ EntrezGene 53904
- ^ Sellers, James R. (2000). "Myosins: a Diverse Superfamily". Biochimica et Biophysica Acta 1496 (1): 3–22. doi:10.1016/S0167-4889(00)00005-7. PMID 10722873.
- ^ =Menetrey, J., Llinas, P., Mukherjea, M., Sweeney, H.L., Houdusse, A. (2007). "The structural basis for the large powerstroke of myosin VI.". Cell 131: 300–8. doi:10.1016/j.cell.2007.08.027. PMID 17956731.
- ^ Buss, Folma; Kendrick-Jones, John (2008). "How are the cellular functions of myosin VI regulated within the cell?". Biochemical and Biophysical Research Communications 369 (1): 165–175. doi:10.1016/j.bbrc.2007.11.150. PMC 2635068. PMID 18068125. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2635068.
- ^ Buss, Folma; Spudich, Giulietta; Kendrick-Jones, John (2004). "MYOSIN VI: Cellular Functions and Motor Properties". Annual Review of Cell and Developmental Biology 20: 649–76. doi:10.1146/annurev.cellbio.20.012103.094243. PMID 15473855.
- ^ Schliwa, Manfred, ed (2003). Molecular Motors. Wiley-VCH. pp. 516, 518. ISBN 3-527-30594-7.
- ^ Reddy, Anireddy S. N.; Day, Irene S. (2001). "Analysis of the myosins encoded in the recently completed Arabidopsis thaliana genome sequence". Genome biology 2 (7): RESEARCH0024. PMC 55321. PMID 11516337. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=55321.
- ^ Baluka, František; Cvrčková, Fatima; Kendrick-Jones, John; Volkmann, Dieter (2001). "Sink Plasmodesmata as Gateways for Phloem Unloading. Myosin VIII and Calreticulin as Molecular Determinants of Sink Strength?". Plant physiology 126 (1): 39–46. doi:10.1104/pp.126.1.39. PMC 1540107. PMID 11351069. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1540107.
- ^ Reichelt, Stefanie; Knight, Alex E.; Hodge, Tony P.; Baluska, Frantisek; Samaj, Jozef; Volkmann, Dieter; Kendrick-Jones, John (1999). "Characterization of the unconventional myosin VIII in plant cells and its localization at the post-cytokinetic cell wall". The Plant journal 19 (5): 555–67. doi:10.1046/j.1365-313X.1999.00553.x. PMID 10504577.
- ^ Inoue, Akira; Saito, Junya; Ikebe, Reiko; Ikebe, Mitsuo (2002). "Myosin IXb is a single-headed minus-end-directed processive motor". Nature cell biology 4 (4): 302–6. doi:10.1038/ncb774. PMID 11901422.
- ^ Sattarzadeh, A.; Krahmer, J.; Germain, A. D.; Hanson, M. R. (2009). "A Myosin XI Tail Domain Homologous to the Yeast Myosin Vacuole-Binding Domain Interacts with Plastids and Stromules in Nicotiana benthamiana". Molecular Plant 2 (6): 1351–8. doi:10.1093/mp/ssp094. PMID 19995734.
- ^ Delbac, F.; Sänger, A; Neuhaus, EM; Stratmann, R; Ajioka, JW; Toursel, C; Herm-Götz, A; Tomavo, S et al. (2001). "Toxoplasma gondii myosins B/C: one gene, two tails, two localizations, and a role in parasite division". The Journal of Cell Biology 155 (4): 613–23. doi:10.1083/jcb.200012116. PMC 2198869. PMID 11706051. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2198869.
- ^ Hettmann, Christine; Herm, Angelika; Geiter, Ariane; Frank, Bernd; Schwarz, Eva; Soldati, Thierry; Soldati, Dominique (2000). "A Dibasic Motif in the Tail of a Class XIV Apicomplexan Myosin Is an Essential Determinant of Plasma Membrane Localization". Molecular biology of the cell 11 (4): 1385–400. PMC 14854. PMID 10749937. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=14854.
- ^ a b Winkelman, L (1976). "Comparative studies of paramyosins". Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 55 (3): 391–7. doi:10.1016/0305-0491(76)90310-2.
- ^ Twarog, B. M. (1976). "Aspects of smooth muscle function in molluscan catch muscle". Physiological reviews 56 (4): 829–38. PMID 185635.
Further reading
- Gavin, R.H. (2001). "Myosins in protists". A Survey of Cell Biology. International Review of Cytology. 206. pp. 97–134. doi:10.1016/S0074-7696(01)06020-X. ISBN 9780123646101.
- Mooseker, M S; Cheney, R E (1995). "Unconventional Myosins". Annual Review of Cell and Developmental Biology 11: 633–75. doi:10.1146/annurev.cb.11.110195.003221. PMID 8689571.
- Sellers, J (2000). "Myosins: a diverse superfamily". Biochimica et Biophysica Acta 1496 (1): 3–22. doi:10.1016/S0167-4889(00)00005-7. PMID 10722873.
- Soldati, Thierry; Geissler, Heidrun; Schwarz, Eva C. (1999). "How many is enough? exploring the myosin repertoire in the model eukaryoteDictyostelium discoideum". Cell Biochemistry and Biophysics 30 (3): 389–411. doi:10.1007/BF02738121. PMID 10403058.
- Molecular Biology of the Cell. Alberts, Johnson, Lewis, Raff, Roberts, and Walter. 4th Edition. 949-952.
Additional images
-

Phase 1
-

Phase 2
-

Phase 3
-

Phase 4
External links
- Myosin Video A video of a moving myosin motor protein.
- MeSH Myosins
- The Myosin Homepage
- http://cellimages.ascb.org/cdm4/item_viewer.php?CISOROOT=/p4041coll12&CISOPTR=101&CISOBOX=1&REC=2[dead link] Animation of a moving myosin motor protein
- EC 3.6.4.1
Proteins of the cytoskeleton Human MyosinsI (MYO1A, MYO1B, MYO1C, MYO1D, MYO1E, MYO1F, MYO1G, MYO1H) · II (MYH1, MYH2, MYH3, MYH4, MYH6, MYH7, MYH7B, MYH8, MYH9, MYH10, MYH11, MYH13, MYH14, MYH15, MYH16) · III (MYO3A, MYO3B) · V (MYO5A, MYO5B, MYO5C) · VI (MYO6) · VII (MYO7A, MYO7B) · IX (MYO9A, MYO9B) · X (MYO10) · XV (MYO15A) · XVIII (MYO18A, MYO18B) · LC (MYL1, MYL2, MYL3, MYL4, MYL5, MYL6, MYL6B, MYL7, MYL9, MYLIP, MYLK, MYLK2, MYLL1)OtherOtherEpithelial keratins
(soft alpha-keratins)Hair keratins
(hard alpha-keratins)Ungrouped alphaNot alphaType 3Type 4Type 5OtherOtherNonhuman Hydrolases: acid anhydride hydrolases (EC 3.6) 3.6.1 3.6.2 3.6.3-4: ATPase 3.6.3Cu++ (3.6.3.4)Ca+ (3.6.3.8)Na+/K+ (3.6.3.9)H+/K+ (3.6.3.10)ATP4AOther P-type ATPase3.6.43.6.5: GTPase 3.6.5.1: Heterotrimeric G protein3.6.5.2: Small GTPase > Ras superfamily3.6.5.3: Protein-synthesizing GTPase3.6.5.5-6: Polymerization motorsHistology: muscle tissue (TH H2.00.05, H3.3) Smooth
muscleStriated
muscleCostamere/
DAPCMembrane/
extracellularIntracellularDystrophin · Dystrobrevin (A, B) · Syntrophin (A, B1, B2, G1, G2) · Syncoilin · Dysbindin · Synemin/desmuslin
related: NOS1 · Caveolin 3Myofilament (thin filament/actin, thick filament/myosin, elastic filament/titin, nebulin)
Troponin (T, C, I)GeneralNeuromuscular junction · Motor unit · Muscle spindle · Excitation-contraction coupling · Sliding filament mechanismBothFiberCellsOtherOther/
ungroupedCategories:- Proteins
- Motor proteins
- Cytoskeleton
- Skeletal muscle







Wikimedia Foundation. 2010.