- Nucleosome
-
Nucleosomes are the basic unit of DNA packaging in eukaryotes, consisting of a segment of DNA wound around a histone protein core.[1] This structure is often compared to thread wrapped around a spool.[2]
Nucleosomes form the fundamental repeating units of eukaryotic chromatin,[3] which is used to pack the large eukaryotic genomes into the nucleus while still ensuring appropriate access to it (in mammalian cells approximately 2 m of linear DNA have to be packed into a nucleus of roughly 10 µm diameter). Nucleosomes are folded through a series of successively higher order structures to eventually form a chromosome; this both compacts DNA and creates an added layer of regulatory control, which ensures correct gene expression. Nucleosomes are thought to carry epigenetically inherited information in the form of covalent modifications of their core histones. The nucleosome hypothesis was proposed by Don and Ada Olins in 1974[4] and Roger Kornberg.[5][6]
The nucleosome core particle consists of approximately 147[7] base pairs of DNA wrapped in 1.67 left-handed superhelical turns around a histone octamer consisting of 2 copies each of the core histones H2A, H2B, H3, and H4.[8] Core particles are connected by stretches of "linker DNA", which can be up to about 80 bp long. Technically, a nucleosome is defined as the core particle plus one of these linker regions; however the word is often synonymous with the core particle.[9]
Linker histones such as H1 and its isoforms are involved in chromatin compaction and sit at the base of the nucleosome near the DNA entry and exit binding to the linker region of the DNA.[10] Non-condensed nucleosomes without the linker histone resemble "beads on a string of DNA" under an electron microscope.[11]
In contrast to most eukaryotic cells, mature sperm cells largely use protamines to package their genomic DNA, most likely to achieve an even higher packaging ratio.[12] Histone equivalents and a simplified chromatin structure have also been found in Archea,[13] proving that eukaryotes are not the only organisms that use nucleosomes.
Contents
Structure
Structure of the core particle
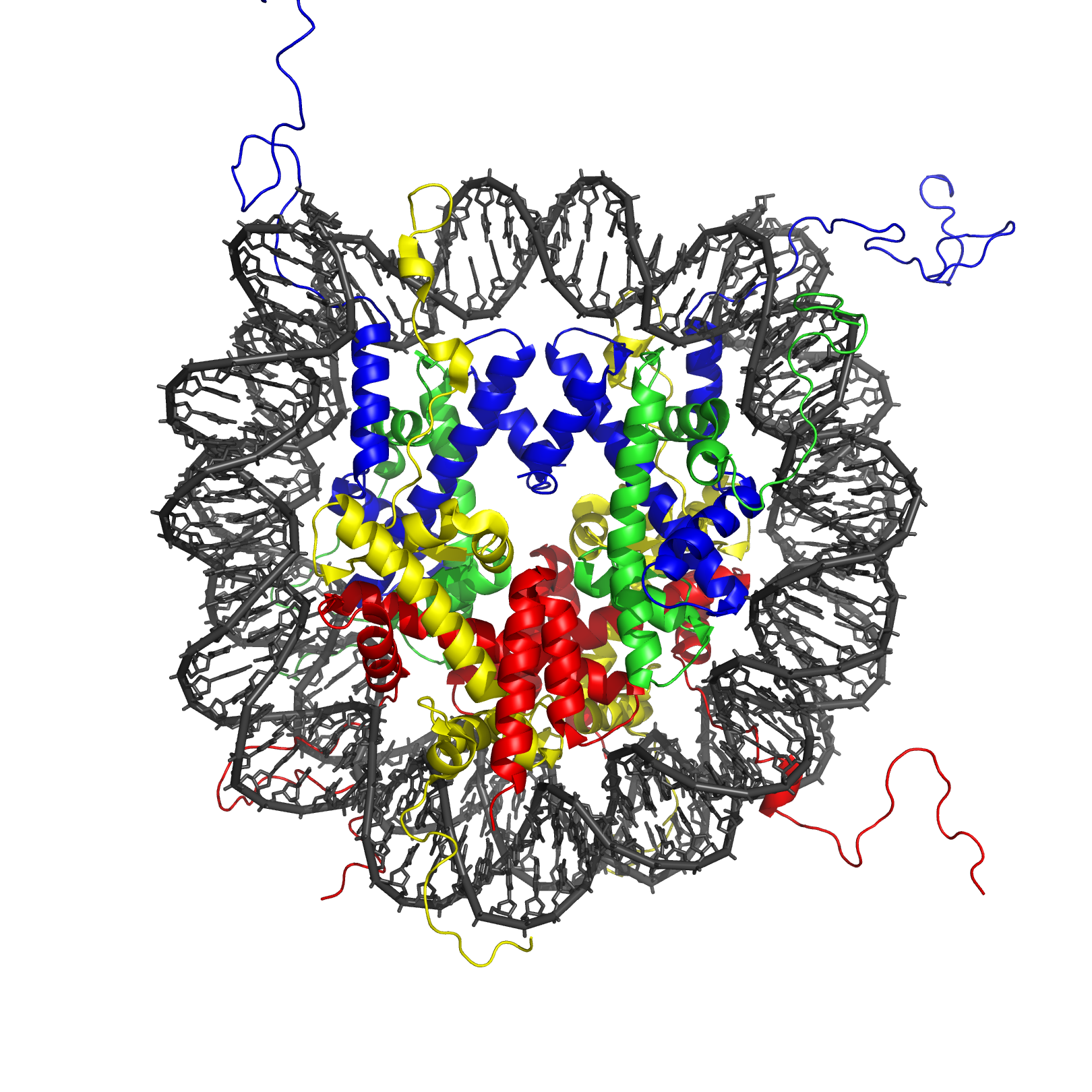
 The crystal structure of the nucleosome core particle consisting of H2A , H2B , H3 and H4 core histones, and DNA. The view is from the top through the superhelical axis.
The crystal structure of the nucleosome core particle consisting of H2A , H2B , H3 and H4 core histones, and DNA. The view is from the top through the superhelical axis.
Overview
Early structural studies provided evidence that an octamer of histone proteins wraps DNA around itself in about two turns of a left-handed superhelix. In 1997 the first near atomic resolution crystal structure of the nucleosome was solved by the Richmond group, showing some of the most important details of the particle. The human alpha-satellite palindromic DNA critical to achieving the 1997 nucleosome crystal structure was developed by the Bunick group at Oak Ridge National Laboratory in Tennessee.[14][15][16][17][18] The structures of over 20 different nucleosome core particles have been solved to date,[19] including those containing histone variants and histones from different species. The structure of the nucleosome core particle is remarkably conserved, and even a change of over 100 residues between frog and yeast histones results in electron density maps with an overall root mean square deviation of only 1.6Å.[20]
The nucleosome core particle
The nucleosome core particle (shown in the figure) consists of about 146[7] bp of DNA wrapped in 1.67 left-handed superhelical turns around the histone octamer, consisting of 2 copies each of the core histones H2A, H2B, H3, and H4. Adjacent nucleosomes are joined by a stretch of free DNA termed "linker DNA" (which varies from 10 - 80 bp in length depending on species and tissue type[13]).
Nucleosome core particles are observed when chromatin in interphase is treated to cause the chromatin to unfold partially. The resulting image, via an electron microscope, is "beads on a string". The string is the DNA, while each bead in the nucleosome is a core particle. The nucleosome core particle is composed of DNA and histone proteins.[21]
Protein interactions within the nucleosome
The core histone proteins contain a characteristic structural motif termed the "histone fold," which consists of three alpha-helices (α1-3) separated by two loops (L1-2). In solution, the histones form H2A-H2B heterodimers and H3-H4 heterotetramers. Histones dimerise about their long α2 helices in an anti-parallel orientation, and, in the case of H3 and H4, two such dimers form a 4-helix bundle stabilised by extensive H3-H3’ interaction. The H2A/H2B dimer binds onto the H3/H4 tetramer due to interactions between H4 and H2B, which include the formation of a hydrophobic cluster.[8] The histone octamer is formed by a central H3/H4 tetramer sandwiched between two H2A/H2B dimers. Due to the highly basic charge of all four core histones, the histone octamer is stable only in the presence of DNA or very high salt concentrations.
Histone - DNA interactions
The nucleosome contains over 120 direct protein-DNA interactions and several hundred water-mediated ones.[22] Direct protein - DNA interactions are not spread evenly about the octamer surface but rather located at discrete sites. These are due to the formation of two types of DNA binding sites within the octamer; the α1α1 site, which uses the α1 helix from two adjacent histones, and the L1L2 site formed by the L1 and L2 loops. Salt links and hydrogen bonding between both side-chain basic and hydroxyl groups and main-chain amides with the DNA backbone phosphates form the bulk of interactions with the DNA. This is important, given that the ubiquitous distribution of nucleosomes along genomes requires it to be a non-sequence-specific DNA-binding factor. Although nucleosomes tend to prefer some DNA sequences over others,[23] they are capable of binding practically to any sequence, which is thought to be due to the flexibility in the formation of these water-mediated interactions. In addition, non-polar interactions are made between protein side-chains and the deoxyribose groups, and an arginine side-chain intercalates into the DNA minor groove at all 14 sites it faces the octamer surface. The distribution and strength of DNA-binding sites about the octamer surface distorts the DNA within the nucleosome core. The DNA is non-uniformly bent and also contains twist defects. The twist of free B-form DNA in solution is 10.5 bp per turn. However, the overall twist of nucleosomal DNA is only 10.2 bp per turn, varying from a value of 9.4 to 10.9 bp per turn.
Histone tail domains
The histone tail extensions constitute up to 30% by mass of histones, but are not visible in the crystal structures of nucleosomes due to their high intrinsic flexibility, and have been thought to be largely unstructured.[24] The N-terminal tails of histones H3 and H2B pass through a channel formed by the minor grooves of the two DNA strands, protruding from the DNA every 20 bp. The N-terminal tail of histone H4, on the other hand, has a region of highly basic amino acids (16-25), which, in the crystal structure, forms an interaction with the highly acidic surface region of a H2A-H2B dimer of another nucleosome, being potentially relevant for the higher-order structure of nucleosomes. This interaction is thought to occur under physiological conditions also, and suggests that acetylation of the H4 tail distorts the higher-order structure of chromatin.
Higher order structure
 The current chromatin compaction model.
The current chromatin compaction model.The organization of the DNA that is achieved by the nucleosome cannot fully explain the packaging of DNA observed in the cell nucleus. Further compaction of chromatin into the cell nucleus is necessary, but is not yet well understood. The current understanding[19] is that repeating nucleosomes with intervening "linker" DNA form a 10-nm-fiber, described as "beads on a string", and have a packing ratio of about five to ten.[13] A chain of nucleosomes can be arranged in a 30 nm fiber, a compacted structure with a packing ratio of ~50[13] and whose formation is dependent on the presence of the H1 histone.
A crystal structure of a tetranucleosome has been presented and used to build up a proposed structure of the 30 nm fiber as a two-start helix.[25] There is still a certain amount of contention regarding this model, as it is incompatible with recent electron microscopy data.[26] Beyond this, the structure of chromatin is poorly understood, but it is classically suggested that the 30 nm fiber is arranged into loops along a central protein scaffold to form transcriptionally active euchromatin. Further compaction leads to transcriptionally inactive heterochromatin.
Nucleosome dynamics
Although the nucleosome is a very stable protein-DNA complex, it is not static and has been shown to undergo a number of different structural re-arrangements including nucleosome sliding and DNA site exposure. Depending on the context, nucleosomes can inhibit or facilitate transcription factor binding. Nucleosome positions are controlled by three major contributions: First, the intrinsic binding affinity of the histone octamer depends on the DNA sequence. Second, the nucleosome can be displaced or recruited by the competitive or cooperative binding of other protein factors. Third, the nucleosome may be actively translocated by ATP-dependent remodeling complexes.[27]
Nucleosome sliding
Work performed in the Bradbury laboratory showed that nucleosomes reconstituted onto the 5S DNA positioning sequence were able to reposition themselves translationally onto adjacent sequences when incubated thermally.[28] Later work showed that this repositioning did not require disruption of the histone octamer but was consistent with nucleosomes being able to “slide” along the DNA in cis. In 2008, It was further revealed that CTCF binding sites act as nucleosome positioning anchors so that, when used to align various genomic signals, multiple flanking nucleosomes can be readily identified.[29] Although nucleosomes are intrinsically mobile, eukaryotes have evolved a large family of ATP-dependent chromatin remodelling enzymes to alter chromatin structure, many of which do so via nucleosome sliding.
DNA site exposure
Work from the Widom laboratory has shown that nucleosomal DNA is in equilibrium between a wrapped and unwrapped state. Measurements of these rates using time-resolved FRET revealed that DNA within the nucleosome remains fully wrapped for only 250 ms before it is unwrapped for 10-50 ms and then rapidly rewrapped.[30] This implies that DNA does not need to be actively dissociated from the nucleosome but that there is a significant fraction of time during which it is fully accessible. Indeed, this can be extended to the observation that introducing a DNA-binding sequence within the nucleosome increases the accessibility of adjacent regions of DNA when bound.[31] This propensity for DNA within the nucleosome to “breathe” is predicted to have important functional consequences for all DNA-binding proteins that operate in a chromatin environment.[citation needed]
Modulating nucleosome structure
Eukaryotic genomes are ubiquitously associated into chromatin; however, cells must spatially and temporally regulate specific loci independently of bulk chromatin. In order to achieve the high level of control required to co-ordinate nuclear processes such as DNA replication, repair, and transcription, cells have developed a variety of means to locally and specifically modulate chromatin structure and function. This can involve covalent modification of histones, the incorporation of histone variants, and non-covalent remodelling by ATP-dependent remodelling enzymes.
Histone post-translational modifications
Since they were discovered in the mid-1960s, histone modifications have been predicted to affect transcription.[32] The fact that most of the early post-translational modifications found were concentrated within the tail extensions that protrude from the nucleosome core lead to two main theories regarding the mechanism of histone modification. The first of the theories suggested that they may affect electrostatic interactions between the histone tails and DNA to “loosen” chromatin structure. Later it was proposed that combinations of these modifications may create binding epitopes with which to recruit other proteins.[33] Recently, given that more modifications have been found in the structured regions of histones, it has been put forward that these modifications may affect histone-DNA[34] and histone-histone[35] interactions within the nucleosome core. Some modifications have been shown to be correlated with gene silencing; others seem to be correlated with gene activation. Common modifications include acetylation, methylation, or ubiquitination of lysine; methylation of arginine; and phosphorylation of serine. The information stored in this way is considered epigenetic, since it is not encoded in the DNA but is still inherited to daughter cells. The maintenance of a repressed or activated status of a gene is often necessary for cellular differentiation.[13]
Histone variants
Although histones are remarkably conserved throughout evolution, several variant forms have been identified. It is interesting to note that this diversification of histone function is restricted to H2A and H3, with H2B and H4 being mostly invariant. H2A can be replaced by H2AZ (which leads to reduced nucleosome stability) or H2AX (which is associated with DNA repair and T cell differentiation), whereas the inactive X chromosomes in mammals are enriched in macroH2A. H3 can be replaced by H3.3 (which correlates with activated genes) and in centromeres H3 is replaced by CENPA.[13]
ATP-dependent nucleosome remodelling
A number of distinct reactions are associated with the term ATP-dependent chromatin remodelling. Remodelling enzymes have been shown to slide nucleosomes along DNA,[36] disrupt histone-DNA contacts to the extent of destabilising the H2A/H2B dimer[37][38] and to generate negative superhelical torsion in DNA and chromatin.[39] Recently, the Swr1 remodelling enzyme has been shown to introduce the variant histone H2A.Z into nucleosomes.[40] At present, it is not clear if all of these represent distinct reactions or merely alternative outcomes of a common mechanism. What is shared between all, and indeed the hallmark of ATP-dependent chromatin remodelling, is that they all result in altered DNA accessibility. Studies looking at gene activation in vivo[41] and, more astonishingly, remodelling in vitro[42] have revealed that chromatin remodelling events and transcription-factor binding are cyclical and periodic in nature. While the consequences of this for the reaction mechanism of chromatin remodelling are not known, the dynamic nature of the system may allow it to respond faster to external stimuli.
Dynamic nucleosome remodelling across the Yeast genome
Studies in 2007 have catalogued nucleosome positions in yeast and shown that nucleosomes are depleted in promoter regions and origins of replication.[43][44][45] About 80% of the yeast genome appears to be covered by nucleosomes [46] and the pattern of nucleosome positioning clearly relates to DNA regions that regulate transcription, regions that are transcribed and regions that initiate DNA replication.[47] Most recently, a new study examined ‘’dynamic changes’’ in nucleosome repositioning during a global transcriptional reprogramming event to elucidate the effects on nucleosome displacement during genome-wide transcriptional changes in yeast (Saccharomyces cerevisiae).[48] The results suggested that nucleosomes that were localized to promoter regions are displaced in response to stress (like heat shock). In addition, the removal of nucleosomes usually corresponded to transcriptional activation and the replacement of nucleosomes usually corresponded to transcriptional repression, presumably because transcription factor binding sites became more or less accessible, respectively. In general, only one or two nucleosomes were repositioned at the promoter to effect these transcriptional changes. However, even in chromosomal regions that were not associated with transcriptional changes, nucleosome repositioning was observed, suggesting that the covering and uncovering of transcriptional DNA does not necessarily produce a transcriptional event.
Nucleosome assembly in vitro
 Diagram of nucleosome assembly.
Diagram of nucleosome assembly.Nucleosomes can be assembled in vitro by either using purified native or recombinant histones.[49][50] One standard technique of loading the DNA around the histones involves the use of salt dialysis. A reaction consisting of the histone octamers and a naked DNA template can be incubated together at a salt concentration of 2 M. By steadily decreasing the salt concentration, the DNA will equilibrate to a position where it is wrapped around the histone octamers, forming nucleosomes. In appropriate conditions, this reconstitution process allows for the nucleosome positioning affinity of a given sequence to be mapped experimentally.[51]
References
- ^ Reece, Jane; Campbell, Neil (2006). Biology. San Francisco: Benjamin Cummings. ISBN 0-8053-6624-5.
- ^ Backstage with a command performer News Release, Rockefeller University, Feb. 18, 2003
- ^ Alberts, Bruce (2002). Molecular biology of the cell (4th ed.). New York: Garland Science. p. 207. ISBN 0-8153-4072-9. http://www.ncbi.nlm.nih.gov/books/bv.fcgi?highlight=Nucleosome&rid=mboc4.section.608#630.
- ^ Olins AL, Olins DE (January 1974). "Spheroid chromatin units (v bodies)". Science 183 (4122): 330–2. doi:10.1126/science.183.4122.330. PMID 4128918. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=4128918.
- ^ McDonald D, "Milestone 9, (1973-1974) The nucleosome hypothesis: An alternative string theory", Nature Milestones: Gene Expression. (2005) Dec 1; http://www.nature.com/milestones/geneexpression/milestones/articles/milegene09.html
- ^ Kornberg RD (May 1974). "Chromatin structure: a repeating unit of histones and DNA". Science 184 (4139): 868–71. doi:10.1126/science.184.4139.868. PMID 4825889. http://www.sciencemag.org/cgi/pmidlookup?view=long&pmid=4825889.
- ^ a b In different crystals, values of 146 and 147 basepairs were observed
- ^ a b Luger K, Mäder AW, Richmond RK, Sargent DF, Richmond TJ (September 1997). "Crystal structure of the nucleosome core particle at 2.8 A resolution". Nature 389 (6648): 251–60. doi:10.1038/38444. PMID 9305837.
- ^ Alberts, Bruce (2007). Molecular biology of the cell (5th ed.). New York: Garland Science. p. 211. ISBN 978-0-8153-4106-2.
- ^ Zhou YB, Gerchman SE, Ramakrishnan V, Travers A, Muyldermans S (September 1998). "Position and orientation of the globular domain of linker histone H5 on the nucleosome". Nature 395 (6700): 402–5. doi:10.1038/26521. PMID 9759733. http://www.nature.com/nature/journal/v395/n6700/abs/395402a0.html.
- ^ Thoma F, Koller T, Klug A (November 1979). "Involvement of histone H1 in the organization of the nucleosome and of the salt-dependent superstructures of chromatin". J. Cell Biol. 83 (2 Pt 1): 403–27. doi:10.1083/jcb.83.2.403. PMC 2111545. PMID 387806. http://www.jcb.org/cgi/pmidlookup?view=long&pmid=387806.
- ^ Clarke HJ (1992). "Nuclear and chromatin composition of mammalian gametes and early embryos". Biochem. Cell Biol. 70 (10-11): 856–66. doi:10.1139/o92-134. PMID 1297351.
- ^ a b c d e f Felsenfeld G, Groudine M (January 2003). "Controlling the double helix". Nature 421 (6921): 448–53. doi:10.1038/nature01411. PMID 12540921.
- ^ Electrophoresis. 1995 Oct;16(10):1861-4. Preparative separation of nucleosome core particles containing defined-sequence DNA in multiple translational phases. Harp JM, Palmer EL, York MH, Gewiess A, Davis M, Bunick GJ.
- ^ Large-scale production of palindrome DNA fragments. Palmer EL, Gewiess A, Harp JM, York MH, Bunick GJ. Anal Biochem. 1995 Oct 10;231(1):109-14.
- ^ Acta Crystallogr D Biol Crystallogr. 1996 Mar 1;52(Pt 2):283-8. X-ray diffraction analysis of crystals containing twofold symmetric nucleosome core particles. Harp JM, Uberbacher EC, Roberson AE, Palmer EL, Gewiess A, Bunick GJ.
- ^ Acta Crystallogr D Biol Crystallogr. 2000 Dec;56(Pt 12):1513-34. Asymmetries in the nucleosome core particle at 2.5 A resolution. Harp JM, Hanson BL, Timm DE, Bunick GJ.
- ^ Methods Enzymol. 2004;375:44-62. Preparation and crystallization of nucleosome core particle. Hanson BL, Alexander C, Harp JM, Bunick GJ.
- ^ a b Chakravarthy S, Park YJ, Chodaparambil J, Edayathumangalam RS, Luger K (February 2005). "Structure and dynamic properties of nucleosome core particles". FEBS Lett. 579 (4): 895–8. doi:10.1016/j.febslet.2004.11.030. PMID 15680970.
- ^ White CL, Suto RK, Luger K (September 2001). "Structure of the yeast nucleosome core particle reveals fundamental changes in internucleosome interactions". EMBO J. 20 (18): 5207–18. doi:10.1093/emboj/20.18.5207. PMC 125637. PMID 11566884. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=125637.
- ^ Alberts, Bruce. Essential Cell Biology. 2nd ed. New York: Garland Science, 2009. Print.
- ^ Davey CA, Sargent DF, Luger K, Maeder AW, Richmond TJ (June 2002). "Solvent mediated interactions in the structure of the nucleosome core particle at 1.9 a resolution". J. Mol. Biol. 319 (5): 1097–113. doi:10.1016/S0022-2836(02)00386-8. PMID 12079350.
- ^ Segal E, Fondufe-Mittendorf Y, Chen L, et al. (August 2006). "A genomic code for nucleosome positioning". Nature 442 (7104): 772–8. doi:10.1038/nature04979. PMC 2623244. PMID 16862119. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2623244.
- ^ Zheng C, Hayes JJ (April 2003). "Structures and interactions of the core histone tail domains". Biopolymers 68 (4): 539–46. doi:10.1002/bip.10303. PMID 12666178.
- ^ Schalch T, Duda S, Sargent DF, Richmond TJ (July 2005). "X-ray structure of a tetranucleosome and its implications for the chromatin fibre". Nature 436 (7047): 138–41. doi:10.1038/nature03686. PMID 16001076.
- ^ Robinson PJ, Fairall L, Huynh VA, Rhodes D (April 2006). "EM measurements define the dimensions of the "30-nm" chromatin fiber: evidence for a compact, interdigitated structure". Proc. Natl. Acad. Sci. U.S.A. 103 (17): 6506–11. doi:10.1073/pnas.0601212103. PMC 1436021. PMID 16617109. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1436021.
- ^ Teif VB, Rippe K (2009). "Predicting nucleosome positions on the DNA: combining intrinsic sequence preferences and remodeler activities". Nucleic Acids Res 37 (17): 5641–5655. doi:10.1093/nar/gkp610. PMC 2761276. PMID 19625488. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2761276.
- ^ Pennings S, Muyldermans S, Meersseman G, Wyns L (May 1989). "Formation, stability and core histone positioning of nucleosomes reassembled on bent and other nucleosome-derived DNA". J. Mol. Biol. 207 (1): 183–92. doi:10.1016/0022-2836(89)90449-X. PMID 2738923. http://linkinghub.elsevier.com/retrieve/pii/0022-2836(89)90449-X.
- ^ Fu Y, Sinha M, Peterson CL, Weng Z (2008). Van Steensel, Bas. ed. "The insulator binding protein CTCF positions 20 nucleosomes around its binding sites across the human genome". PLoS genetics 4 (7): e1000138. doi:10.1371/journal.pgen.1000138. PMC 2453330. PMID 18654629. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2453330.
- ^ Li G, Levitus M, Bustamante C, Widom J (January 2005). "Rapid spontaneous accessibility of nucleosomal DNA". Nat. Struct. Mol. Biol. 12 (1): 46–53. doi:10.1038/nsmb869. PMID 15580276.
- ^ Li G, Widom J (August 2004). "Nucleosomes facilitate their own invasion". Nat. Struct. Mol. Biol. 11 (8): 763–9. doi:10.1038/nsmb801. PMID 15258568.
- ^ Allfrey VG, Faulkner R, Mirsky AE (May 1964). "Acetylation and methylation of histones and their possible role in the regulation of RNA synthesis". Proc. Natl. Acad. Sci. U.S.A. 51: 786–94. doi:10.1073/pnas.51.5.786. PMC 300163. PMID 14172992. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=300163.
- ^ Strahl BD, Allis CD (January 2000). "The language of covalent histone modifications". Nature 403 (6765): 41–5. doi:10.1038/47412. PMID 10638745.
- ^ Cosgrove MS, Boeke JD, Wolberger C (November 2004). "Regulated nucleosome mobility and the histone code". Nat. Struct. Mol. Biol. 11 (11): 1037–43. doi:10.1038/nsmb851. PMID 15523479.
- ^ Ye J, Ai X, Eugeni EE, et al. (April 2005). "Histone H4 lysine 91 acetylation a core domain modification associated with chromatin assembly". Mol. Cell 18 (1): 123–30. doi:10.1016/j.molcel.2005.02.031. PMC 2855496. PMID 15808514. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2855496.
- ^ Whitehouse I, Flaus A, Cairns BR, White MF, Workman JL, Owen-Hughes T (August 1999). "Nucleosome mobilization catalysed by the yeast SWI/SNF complex". Nature 400 (6746): 784–7. doi:10.1038/23506. PMID 10466730.
- ^ Kassabov SR, Zhang B, Persinger J, Bartholomew B (February 2003). "SWI/SNF unwraps, slides, and rewraps the nucleosome". Mol. Cell 11 (2): 391–403. doi:10.1016/S1097-2765(03)00039-X. PMID 12620227. http://linkinghub.elsevier.com/retrieve/pii/S109727650300039X.
- ^ Bruno M, Flaus A, Stockdale C, Rencurel C, Ferreira H, Owen-Hughes T (December 2003). "Histone H2A/H2B dimer exchange by ATP-dependent chromatin remodeling activities". Mol. Cell 12 (6): 1599–606. doi:10.1016/S1097-2765(03)00499-4. PMID 14690611. http://linkinghub.elsevier.com/retrieve/pii/S1097276503004994.
- ^ Havas K, Flaus A, Phelan M, et al. (December 2000). "Generation of superhelical torsion by ATP-dependent chromatin remodeling activities". Cell 103 (7): 1133–42. doi:10.1016/S0092-8674(00)00215-4. PMID 11163188. http://linkinghub.elsevier.com/retrieve/pii/S0092-8674(00)00215-4.
- ^ Mizuguchi G, Shen X, Landry J, Wu WH, Sen S, Wu C (January 2004). "ATP-driven exchange of histone H2AZ variant catalyzed by SWR1 chromatin remodeling complex". Science 303 (5656): 343–8. doi:10.1126/science.1090701. PMID 14645854.
- ^ Métivier R, Penot G, Hübner MR, et al. (December 2003). "Estrogen receptor-alpha directs ordered, cyclical, and combinatorial recruitment of cofactors on a natural target promoter". Cell 115 (6): 751–63. doi:10.1016/S0092-8674(03)00934-6. PMID 14675539. http://linkinghub.elsevier.com/retrieve/pii/S0092867403009346.
- ^ Nagaich AK, Walker DA, Wolford R, Hager GL (April 2004). "Rapid periodic binding and displacement of the glucocorticoid receptor during chromatin remodeling". Mol. Cell 14 (2): 163–74. doi:10.1016/S1097-2765(04)00178-9. PMID 15099516. http://linkinghub.elsevier.com/retrieve/pii/S1097276504001789.
- ^ Albert I, Mavrich TN, Tomsho LP, Qi J, Zanton SJ, et al. (2007). "Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome". Nature 446 (7135): 572–576. doi:10.1038/nature05632. PMID 17392789.
- ^ Li B, Carey M, Workman JL (2007). "The Role of Chromatin during Transcription". Cell 128 (4): 707–719. doi:10.1016/j.cell.2007.01.015. PMID 17320508.
- ^ Whitehouse I, Rando OJ, Delrow J, Tsukiyama T (2007). "Chromatin remodelling at promoters suppresses antisense transcription". Nature 450 (7172): 1031–1035. doi:10.1038/nature06391. PMID 18075583.
- ^ Lee W, Tillo D, Bray N, Morse RH, Davis RW, Hughes TR, Nislow C (2007). "A high-resolution atlas of nucleosome occupancy in yeast". Nature Genetics 39 (10): 1235–44. doi:10.1038/ng2117. PMID 17873876.
- ^ Eaton ML, Galani K, Kang S, Bell SP, MacAlpine DM (Apr 2010). "Conserved nucleosome positioning defines replication origins". Genes and Development 24 (8): 748–753. doi:10.1101/gad.1913210. PMC 2854390. PMID 20351051. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2854390.
- ^ Shivaswamy S, Bhinge A, Zhao Y, Jones S, Hirst M, et al. (2007). "Dynamic Remodeling of Individual Nucleosomes Across a Eukaryotic Genome in Response to Transcriptional Perturbation". PLoS Biol 6 (3): e65. doi:10.1371/journal.pbio.0060065. PMC 2267817. PMID 18351804. http://scivee.tv/node/5532.
- ^ Hayes JJ, Lee KM (May 1997). "In vitro reconstitution and analysis of mononucleosomes containing defined DNAs and proteins". Methods 12 (1): 2–9. doi:10.1006/meth.1997.0441. PMID 9169189. http://linkinghub.elsevier.com/retrieve/pii/S1046-2023(97)90441-2.
- ^ Dyer PN, Edayathumangalam RS, White CL, et al. (2004). "Reconstitution of nucleosome core particles from recombinant histones and DNA". Meth. Enzymol. 375: 23–44. PMID 14870657.
- ^ Yenidunya A, Davey C, Clark D, Felsenfeld G, Allan J (April 1994). "Nucleosome positioning on chicken and human globin gene promoters in vitro. Novel mapping techniques". J. Mol. Biol. 237 (4): 401–14. doi:10.1006/jmbi.1994.1243. PMID 8151701. http://linkinghub.elsevier.com/retrieve/pii/S0022-2836(84)71243-5.
External links
- Nucleosomes on the group page of Timothy Richmond
- Proteopedia Nucleosomes
- Nucleosome at the PDB
- Dynamic Remodeling of Individual Nucleosomes Across a Eukaryotic Genome in Response to Transcriptional Perturbation
- Nucleosome positioning data and tools online (annotated list, constantly updated)
Genetics: chromosomes General Classification Evolution Structure Categories:


Wikimedia Foundation. 2010.