- Mammary gland
-
Mammary gland in a human female 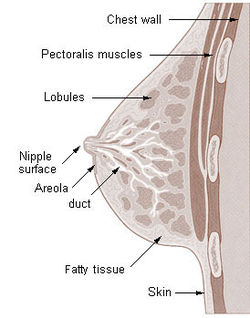
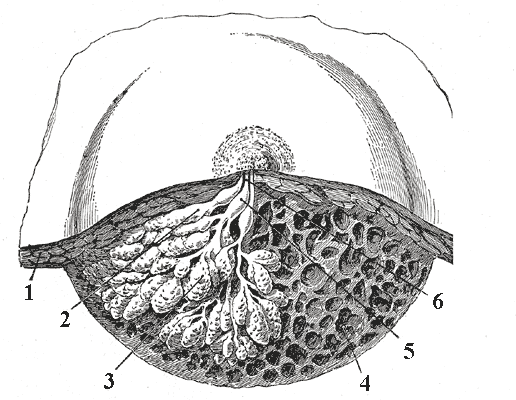
Cross section of the breast of a human female. 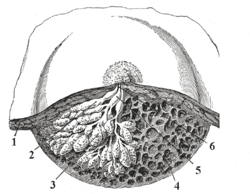
Dissection of a lactating breast.
1 - Fat
2 - Lactiferous duct/lobule
3 - Lobule
4 - Connective tissue
5 - Sinus of lactiferous duct
6 - Lactiferous ductLatin glandula mammaria Gray's subject #271 1267 A mammary gland is an organ in mammals that produces milk to feed young offspring. Mammals get their name from the word "mammary". In ruminants such as cows, goats, and deer, the mammary glands are contained in their udders. The mammary glands of other mammals that have more than two breasts, such as dogs and cats, are sometimes called dugs.
Contents
Humans
Main article: BreastHistology
A mammary gland is a specific type of apocrine gland specialized for manufacture of colostrum at the time of parturition. Mammary glands can be identified as apocrine because they exhibit striking "decapitation" secretion. Whether mammary glands are modified sweat glands or sebaceous glands still remains controversial.[1]
Structure
The basic components of a mature mammary gland are the alveoli (hollow cavities, a few millimetres large) lined with milk-secreting cuboidal cells and surrounded by myoepithelial cells. These alveoli join up to form groups known as lobules, and each lobule has a lactiferous duct that drains into openings in the nipple. The myoepithelial cells can contract under the stimulation of oxytocin thereby excreting milk secreted from alveolar units into the lobule lumen toward the nipple, where it collects in sinuses of the ducts. As the infant begins to suck, the hormonally (oxytocin) mediated "let down reflex" ensues and the mother's milk is secreted – not sucked from the gland – into the baby's mouth.
All the milk-secreting tissue leading to a single lactiferous duct is called a "simple mammary gland"; a "complex mammary gland" is all the simple mammary glands serving one nipple. Humans normally have two complex mammary glands, one in each breast, and each complex mammary gland consists of 10–20 simple glands. The presence of more than two nipples is known as polythelia and the presence of more than two complex mammary glands as polymastia.
To keep the correct polarized morphology of the lactiferous duct tree requires another essential component - mammary epithelial cells extracellular matrix (ECM), which together with adipocytes, fibroblast, inflammatory cells etc. constitute mammary stroma.[2] Mammary epithelial ECM mainly contains myoepithelial basement membrane and the connective tissue. They not only help to support mammary basic structure, but also serve as a communicating bridge between mammary epithelials and their local and global environment throughout this organ's development.[3]
Development and hormonal control
Mammary glands develop during different growth cycles. They exist in both sexes during embryonic stage, forming only a rudimentary duct tree at birth. In this stage, mammary gland development depends on systemic (and maternal) hormones,[2] but is also under the (local) regulation of paracrine communication between neighboring epithelial and mesenchymal cells by parathyroid hormone-related protein(PTHrP).[4] This locally secreted factor gives rise to a series of outside-in and inside-out positive feedback between these two types of cells, so that mammary bud epithelial cells can get to proliferate and sprout down into the mesenchymal layer until they reach the fat pad to begin the first round of branching.[2] At the same time, the embryonic mesenchymal cells around the epithelial bud get secrecting factors activated by PTHrP, such as BMP4, can transform into a dense, mammary-specific mesenchyme, which later develop into connective tissue with fibrous threads, forming blood vessels and the lymph system.[5] Basement membrane, mainly containing laminin and collagen, formed thereafter by differentiated myoepithelial cells keeps the polarity of this primary duct tree.
Lactiferous duct development occurs in females in response to circulating hormones, a first development is frequently seen during pre- and postnatal stages and later during puberty. Estrogen promotes branching differentiation,[6] whereas in males testosterone inhibits it. A mature duct tree reaching the limit of the fat pad of the mammary gland comes into being by bifurcation of duct terminal end buds (TEB), secondary branches sprouting from primary ducts[3][7] and proper duct lumen formation. These processes are tightly modulated by components of mammary epithelial ECM interacting with systemic hormones and local secreting factors. However, for each mechanism the epithelial cells' "niche" can be delicately unique with different membrane receptor profiles and basement membrane thickness from specific branching area to area, so as to regulate cell growth or differentiation sub-locally.[8] Important players include beta-1 integrin, epidermal growth factor receptor (EGFR), laminin-1/5, collagen-IV, matrix metalloproteinase(MMPs), heparan sulfate proteoglycans etc. Elevated circulating level of growth hormone and estrogen get to multipotent cap cells on tip of TEB through a leaky thin layer of basement membrance and promote specific gene expression. Hence cap cells can differentiate into myoepithelial and luminal (duct) epithelial cells, and the increased amount of activated MMPs can degrade surrounding ECM helping duct buds to reach further in the fat pads.[9][10] On the other hand, basement membrane along the mature mammary ducts is thicker with strong adhesion to epithelial cells via binding to integrin and non-integrin receptors. When side branches develop, it is a much more “pushing-forward” working process including extending through myoepithelial cells, degrading basement membrane and then invading into a periductal layer of fibrous stromal tissue.[3] Degraded basement membrane fragments (laminin-5) roles to lead the way of mammary epithelial cells migration.[11] Whereas, laminin-1 interacts with non-integrin receptor dystroglycan negatively regulates this side branching process in case of cancer.[12] These complex "Yin-yang" balancing crosstalks between mammary ECM and epithelial cells "instruct" healthy mammary gland development until adult.
Secretory alveoli develop mainly in pregnancy, when rising levels of prolactin, estrogen and progesterone cause further branching, together with an increase in adipose tissue and a richer blood flow. In gestation, serum progesterone remains at a stably high concentration so signaling through its receptor is continuously activated. As one of the transcribed genes, Wnts secreted from mammary epithelial cells act paracrinely to induce more neighboring cells branching.[13][14] When the lactiferous duct tree is almost ready, "leaves" alveoli are differentiated from luminal epithelial cells and added at the end of each branch. In late pregnancy and for the first few days after giving birth, colostrum is secreted. Milk secretion (lactation) begins a few days later due to reduction in circulating progesterone and the presence of another important hormone prolactin, which mediates further alveologenesis, milk protein production, and regulates osmotic balance and tight junction function. Laminin and collagen in myoepithelial basement membrane interacting with beta-1 integrin on epithelial surface again, is essential in this process.[15][16] Their binding ensures correct placement of prolactin receptors on basal lateral side of alveoli cells and directional secretion of milk into lactiferous ducts.[15][16] Suckling of the baby causes release of hormone oxytocin which stimulates contraction of the myoepithelial cells. In this way of combined control from ECM and systemic hormones, milk secretion can be reciprocally amplified so as to provide enough nutrition for the baby.
During weaning, decreased prolactin, missing mechanical stimulation (baby suckling) and changes in osmotic balance caused by milk stasis and leaking of tight junctions cause cessation of milk production. In some species there is complete or partial involution of alveolar structures after weaning, in humans there is only partial involution and the level of involution in humans appears to be highly individual. In some other species (such as cows) all alveoli and secretory duct structure collapse by programmed cell death (apoptosis) and autophagy for lack of growth promoting factors either from the ECM or circulating hormones.[17][18] At the same time, apoptosis of blood capillary endothelial cells speeds up the regression of lactation ductal beds. Shrinkage of the mammary duct tree and ECM remodeling by various proteinase is under the control of somatostatin and other growth inhibiting hormones and local factors.[19] This big structure change leads loose fat tissue to fill up the empty space thereafter. But a functional lactiferous duct tree can be formed again when a female is pregnant again.
Breast cancer
Tumorigenesis in mammary glands can be induced biochemically by abnormal expression level of circulating hormones or local ECM components,[20] or from a mechanical change in the tension of mammary stroma.[21] Under either of the two circumstances, mammary epithelial cells would grow out of control and eventually result in cancer. Almost all instances of breast cancer originate in the lobules or ducts of the mammary glands.
Other mammals
The constantly protruding breasts of the adult human female, unusually large relative to body size, are a unique evolutionary development whose purpose is not yet fully known (see breasts); other mammals tend to have less conspicuous mammary glands that protrude only while actually filling with milk. The number and positioning of complex and simple mammary glands varies widely in different mammals. The nipples and glands can occur anywhere along the two milk lines, two roughly-parallel lines along the ventral aspect of the body. In general most mammals develop mammary glands in pairs along these lines, with a number approximating the number of young typically birthed at a time. The number of nipples varies from 2 (in most primates) to 18 (in pigs). The Virginia Opossum has 13, one of the few mammals with an odd number.[22][23] The following table lists the number and position of glands normally found in a range of mammals:
Species [24] Anterior
(thoracic)Intermediate
(abdominal)Posterior
(inguinal)Total Goat, sheep, horse
guinea pig0 0 2 2 Cattle 0 0 4 4 Cat 2 2 4 8 Dog [25] 4 2 2 or 4 8 or 10 Mouse 6 0 4 10 Rat 6 2 4 12 Pig 6 6 6 18 proboscideans, primates 2 0 0 2 Male mammals typically have rudimentary mammary glands and nipples, with a few exceptions: male mice don't have nipples, and male horses lack nipples and mammary glands.[citation needed] The male Dayak fruit bat has lactating mammary glands;[26] male lactation occurs infrequently in some species, including humans.
Mammary glands are true protein factories, and several companies[who?] have constructed transgenic animals, mainly goats and cows, in order to produce proteins for pharmaceutical use. Complex glycoproteins such as monoclonal antibodies or antithrombin cannot be produced by genetically engineered bacteria, and the production in live mammals is much cheaper than the use of mammalian cell cultures.
Evolution
It is believed that the mammary gland is a transformed sweat gland, more closely related to Apocrine sweat glands.[27] There are many theories of how they evolved, but since they do not fossilize well, supporting such theories is difficult. Many of the current theories are based on comparisons between lines of living mammals- monotremes, marsupials and eutherians. One theory proposes that mammary glands evolved from glands that were used to keep the eggs of early mammals moist[28][29] and free from infection[30][31] (monotremes still lay eggs). Other theories propose that early secretions were used directly by hatched young,[32] or that the secretions were used by young to help them orient to their mothers.[33]
Lactation developed long before the evolution of the mammary gland and mammals, see evolution of lactation.
Gallery
See also
References
- ^ Ackerman, A. Bernard; Almut Böer, Bruce Bennin, and Geoffrey J. Gottlieb (2005). Histologic Diagnosis of Inflammatory Skin Diseases An Algorithmic Method Based on Pattern Analysis. ISBN 978-1893357259. http://www.derm101.com/content/13501.
- ^ a b c Watson, C.J. & Khaled, W.T. Mammary development in the embryo and adult: a journey of morphogenesis and commitment. Development 135, 995-1003 (2008)
- ^ a b c Wiseman, B.S. & Werb, Z. Stromal effects on mammary gland development and breast cancer. Science 296, 1046-9 (2002)
- ^ Wysolmerski, J.J. et al. Rescue of the parathyroid hormone-related protein knockout mouse demonstrates that parathyroid hormone-related protein is essential for mammary gland development. Development 125, 1285-94 (1998)
- ^ Hens, J.R. & Wysolmerski, J.J. Key stages of mammary gland development: molecular mechanisms involved in the formation of the embryonic mammary gland. Breast Cancer Res 7, 220-4 (2005)
- ^ Sternlicht, M.D. Key stages in mammary gland development: the cues that regulate ductal branching morphogenesis. Breast Cancer Res 8, 201 (2006)
- ^ Sternlicht, M.D., Kouros-Mehr, H., Lu, P. & Werb, Z. Hormonal and local control of mammary branching morphogenesis. Differentiation 74, 365-81 (2006)
- ^ Fata, J.E., Werb, Z. & Bissell, M.J. Regulation of mammary gland branching morphogenesis by the extracellular matrix and its remodeling enzymes. Breast Cancer Res 6, 1-11 (2004).
- ^ Wiseman, B.S. et al. Site-specific inductive and inhibitory activities of MMP-2 and MMP-3 orchestrate mammary gland branching morphogenesis. J Cell Biol 162, 1123-33 (2003)
- ^ Koshikawa, N., Giannelli, G., Cirulli, V., Miyazaki, K. & Quaranta, V. Role of cell surface metalloprotease MT1-MMP in epithelial cell migration over laminin-5. J Cell Biol 148, 615-24 (2000)
- ^ Dogic, D., Rousselle, P. & Aumailley, M. Cell adhesion to laminin 1 or 5 induces isoform-specific clustering of integrins and other focal adhesion components. J Cell Sci 111 ( Pt 6), 793-802 (1998)
- ^ Muschler, J. et al. A role for dystroglycan in epithelial polarization: loss of function in breast tumor cells. Cancer Res 62, 7102-9 (2002)
- ^ Robinson, G.W., Hennighausen, L. & Johnson, P.F. Side-branching in the mammary gland: the progesterone-Wnt connection. Genes Dev 14, 889-94 (2000)
- ^ Brisken, C. et al. Essential function of Wnt-4 in mammary gland development downstream of progesterone signaling. Genes Dev 14, 650-4 (2000)
- ^ a b Streuli, C.H., Bailey, N. & Bissell, M.J. Control of mammary epithelial differentiation: basement membrane induces tissue-specific gene expression in the absence of cell-cell interaction and morphological polarity. J Cell Biol 115, 1383-95 (1991)
- ^ a b Streuli, C.H. et al. Laminin mediates tissue-specific gene expression in mammary epithelia. J Cell Biol 129, 591-603 (1995)
- ^ Zarzynska J, Motyl T.Apoptosis and autophagy in involuting bovine mammary gland. J Physiol Pharmacol. 2008 Dec;59 Suppl 9:275-88
- ^ Fadok, V.A., Clearance: the last and often forgotten stage of apoptosis. J Mammary Gland Biol Neoplasia, 1999. 4(2): p. 203-11
- ^ Motyl, T., et al., Apoptosis and autophagy in mammary gland remodeling and breast cancer chemotherapy. J Physiol Pharmacol, 2006. 57 Suppl 7: p. 17-32
- ^ Gudjonsson, T. et al. Normal and tumor-derived myoepithelial cells differ in their ability to interact with luminal breast epithelial cells for polarity and basement membrane deposition. J Cell Sci 115, 39-50 (2002)
- ^ Provenzano, P.P. et al. Collagen density promotes mammary tumor initiation and progression. BMC Med 6, 11 (2008)
- ^ With the Wild Things - Transcripts
- ^ Raising Orphaned Baby Opossums
- ^ Merle Cunningham, Animal Science and Industry ISBN 9780130462565
- ^ Dog breeds vary in the number of mammary glands: larger breeds tend to have 5 pairs, smaller breeds have 4 pairs.
- ^ Francis, Charles M.; Anthony, Edythe L. P.; Brunton, Jennifer A.; Kunz, Thomas H. (24 February 1994). "Lactation in male fruit bats". Nature (Nature Publishing Group) 367 (6465): 691–692. doi:10.1038/367691a0. http://www.nature.com/nature/journal/v367/n6465/abs/367691a0.html. Retrieved 2008-05-14.
- ^ Oftedal, O.T. (2002). "The origin of lactation as a water source for parchment-shelled eggs=Journal of Mammary Gland Biology and Neoplasia". Journal of Mammary Gland Biology and Neoplasia 7 (3): 253–266. doi:10.1023/A:1022848632125. PMID 12751890.
- ^ http://nationalzoo.si.edu/ConservationAndScience/SpotlightOnScience/oftedalolav20030714.cfm
- ^ Oftedal, O.T. (2002). "The mammary gland and its origin during synapsid evolution". Journal of Mammary Gland Biology and Neoplasia 7 (3): 225–252. doi:10.1023/A:1022896515287. PMID 12751889.
- ^ http://scienceblogs.com/pharyngula/2006/05/breast_beginnings.php
- ^ Vorbach C., Capecchi M.R., Penninger J.M. (2006). "Evolution of the mammary gland from the innate immune system?". Bioessays 28 (28): 606–616. doi:10.1002/bies.20423. PMID 16700061.
- ^ Lefèvre C.M., Sharp J.A., Nicholas K.R. (2010). "Evolution of lactation: ancient origin and extreme adaptations of the lactation system". Annual Review of Genomics and Human Genetics (11): 219–238.
- ^ Graves B.M and Duvall D. (1983). "A role for aggregation pheremones in the evolution of mammallike reptile lactation". The American Naturalist 122 (6): 835–839. doi:10.1086/284177.
External links
- Comparative Mammary Gland Anatomy by W. L. Hurley
- On the anatomy of the breast by Sir Astley Paston Cooper (1840). Numerous drawings, in the public domain.
- mammary+gland at eMedicine Dictionary
Female breast (TA A16.0.02, GA 11.1267) Nipple/Areola Areolar glands · Crista supraareolaris (only fusiform breast)Mammary glands Forms Mamma fusiformis · Mamma spherica · Mamma conicaOther Categories:- Breast anatomy
- Exocrine system
- Glands









Wikimedia Foundation. 2010.