- Mirror neuron
-
A mirror neuron is a neuron that fires both when an animal acts and when the animal observes the same action performed by another.[1][2] [3] Thus, the neuron "mirrors" the behaviour of the other, as though the observer were itself acting. Such neurons have been directly observed in primate and other species including birds. In humans, brain activity consistent with that of mirror neurons has been found in the premotor cortex, the supplementary motor area, the primary somatosensory cortex and the inferior parietal cortex.
Mirror neurons were first described in 1992. Some scientists consider this to be one of the most important recent discoveries in neuroscience. Among them is V.S. Ramachandran, who believes they might be very important in imitation and language acquisition.[4] However, despite the excitement generated by these findings, to date no widely accepted neural or computational models have been put forward to describe how mirror neuron activity supports cognitive functions such as imitation.[5]
The function of the mirror system is a subject of much speculation. Many researchers in cognitive neuroscience and cognitive psychology consider that this system provides the physiological mechanism for the perception action coupling (see the common coding theory)[3]. These mirror neurons may be important for understanding the actions of other people, and for learning new skills by imitation. Some researchers also speculate that mirror systems may simulate observed actions, and thus contribute to theory of mind skills,[6][7] while others relate mirror neurons to language abilities.[8] It has also been proposed that problems with the mirror system may underlie cognitive disorders, particularly autism.[9][10] However the connection between mirror neuron dysfunction and autism is tentative and it remains to be seen how mirror neurons may be related to many of the important characteristics of autism.[5]
Contents
Discovery
In the 1980s and 1990s, Giacomo Rizzolatti was working with Giuseppe Di Pellegrino, Luciano Fadiga, Leonardo Fogassi, and Vittorio Gallese at the University of Parma, Italy. These neurophysiologists had placed electrodes in the ventral premotor cortex of the macaque monkey to study neurons specialized for the control of hand and mouth actions; for example, taking hold of an object and manipulating it. During each experiment, they recorded from a single neuron in the monkey's brain while the monkey was allowed to reach for pieces of food, so the researchers could measure the neuron's response to certain movements.[11][12] They found that some of the neurons they recorded from would respond when the monkey saw a person pick up a piece of food as well as when the monkey picked up the food. The discovery was initially sent to Nature but was rejected for its “lack of general interest”.[13]
A few years later, the same group published another empirical paper and discussed the role of the mirror neuron system in action recognition, and proposed that the human Broca’s region was the homologue region of the monkey ventral premotor cortex.[14] While these papers reported the presence of mirror neurons responding to hand actions, a subsequent study by Ferrari Pier Francesco and colleagues [15] described the presence of mirror neurons responding to mouth actions and facial gestures.
Further experiments confirmed that about 10% of neurons in the monkey inferior frontal and inferior parietal cortex have 'mirror' properties and give similar responses to performed hand actions and observed actions. More recently Christian Keysers and colleagues have shown that, in both humans and monkeys, the mirror system also responds to the sound of actions.[16][17] [3]
Reports on mirror neurons have been widely published[18] and confirmed[19] with mirror neurons found in both inferior frontal and inferior parietal regions of the brain. Recently, evidence from functional neuroimaging strongly suggests that humans have similar mirror neurons systems: researchers have identified brain regions which respond during both action and observation of action. Not surprisingly, these brain regions include those found in the macaque monkey.[1] However, functional magnetic resonance imaging (fMRI) can examine the entire brain at once and suggests that a much wider network of brain areas shows mirror properties in humans than previously thought. These additional areas include the somatosensory cortex and are thought to make the observer feel what it feels like to move in the observed way [20][21]
In monkeys
 Neonatal (newborn) macaque imitating facial expressions
Neonatal (newborn) macaque imitating facial expressions
The first animal in which mirror neurons have been studied individually is the macaque monkey. In these monkeys, mirror neurons are found in the inferior frontal gyrus (region F5) and the inferior parietal lobule.[1]
Mirror neurons are believed to mediate the understanding of other animals' behaviour. For example, a mirror neuron which fires when the monkey rips a piece of paper would also fire when the monkey sees a person rip paper, or hears paper ripping (without visual cues). These properties have led researchers to believe that mirror neurons encode abstract concepts of actions like 'ripping paper', whether the action is performed by the monkey or another animal.[1]
The function of mirror neurons in macaques is not known. Adult macaques do not seem to learn by imitation. Recent experiments by Ferrari and colleagues suggest that infant macaqes can imitate a human's face movements, though only as neonates and during a limited temporal window.[22] Even if it has not yet been empirically demonstrated, it has been proposed that mirror neurons underlie this behaviour and other imitative phenomena [23].
In adult monkeys, mirror neurons may enable the monkey to understand what another monkey is doing, or to recognise the other monkey's action.[24]
In humans
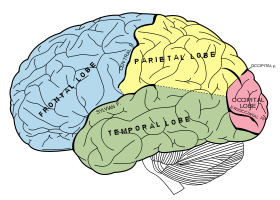 Diagram of the brain, showing the locations of the frontal and parietal lobes of the cerebrum, viewed from the left. The inferior frontal lobe is the lower part of the blue area, and the superior parietal lobe is the upper part of the yellow area.
Diagram of the brain, showing the locations of the frontal and parietal lobes of the cerebrum, viewed from the left. The inferior frontal lobe is the lower part of the blue area, and the superior parietal lobe is the upper part of the yellow area.It is not normally possible to study single neurons in the human brain, so most evidence for mirror neurons in humans is indirect. Brain imaging experiments using functional magnetic resonance imaging (fMRI) have shown that the human inferior frontal cortex and superior parietal lobe is active when the person performs an action and also when the person sees another individual performing an action. It has been suggested that these brain regions contain mirror neurons, and they have been defined as the human mirror neuron system.[25] More recent experiments have shown that even at the level of single participants, scanned using fMRI, large areas containing multiple fMRI voxels increase their activity both during the observation and execution of actions.[20]
Neuropsychological studies looking at lesion areas that cause action knowledge, pantomime interpretation, and biological motion perception deficits have pointed to a causal link between the integrity of the inferior frontal gyrus and these behaviours.[26][27][28] Transcranial magnetic stimulation studies have confirmed this as well.[29][30] These results indicate the activation in mirror neuron related areas are unlikely to be just epiphenomenal.
A study published in April 2010 reports recordings from single neurons with mirror properties in the human brain.[31] Mukamel et al (Current Biology, 2010) recorded from the brains of 21 patients who were being treated at Ronald Reagan UCLA Medical Center for intractable epilepsy. The patients had been implanted with intracranial depth electrodes to identify seizure foci for potential surgical treatment. Electrode location was based solely on clinical criteria; the researchers, with the patients' consent, used the same electrodes to "piggyback" their research. The experiment included three parts: facial expressions, grasping and a control experiment. Activity from a total of 1,177 neurons in the 21 patients was recorded as the patients both observed and performed grasping actions and facial gestures. In the observation phase, the patients observed various actions presented on a laptop computer. In the activity phase, the subjects were asked to perform an action based on a visually presented word. In the control task, the same words were presented and the patients were instructed not to execute the action. The researchers found a small number of neurons that fired or showed their greatest activity both when the individual performed a task and when they observed a task. Other neurons had anti-mirror properties, that is, they responded when the participant saw an action but were inhibited when the participant performed that action. The mirror neurons found were located in the supplementary motor area and medial temporal cortex (other brain regions were not sampled). For purely practical reasons, these regions are not the same as those in which mirror neurons had been recorded from in the monkey: researchers in Parma were studying the ventral premotor cortex and the associated inferior parietal lobe, two regions in which epilepsy rarely occurs, and hence, single cell recordings in these regions are not usually done in humans. On the other hand, no one has to date looked for mirror neurons in the supplementary motor area or the medial temporal lobe in the monkey. Together, this therefore does not suggest that humans and monkeys have mirror neurons in different locations, but rather than they may have mirror neurons both in the ventral premotor cortex and inferior parietal lobe, where they have been recorded in the monkey, and in the supplementary motor areas and medial temporal lobe, where they have been recorded from in human – especially because detailed human fMRI analyses suggest activity compatible with the presence of mirror neurons in all these regions.[20]
Doubts concerning mirror neurons
One recent review argued that the original analyses were unconvincing because they were based on qualitative descriptions of individual cell properties, and did not take into account the small number of strongly mirror-selective neurons in motor areas.[5] Other reviews argued that the measurements of neuron fire delay seem not to be compatible with standard reaction times,[32] and pointed out that nobody has reported that an interruption of the motor areas in F5 would produce a decrement in action recognition.,[33] although it appears these authors have missed human neuropsychological and TMS studies reporting disruption of these areas do indeed cause action deficits [27][29] without affecting other kinds of perception.[28] It is not clear, according to these reviews, whether mirror neurons really form a distinct class of cells (as opposed to an occasional phenomenon seen in cells that have other functions),[32] and whether mirror activity is a distinct type of response or simply an artifact of an overall facilitation of the motor system.[33] Indeed, there is limited understanding of the degree to which monkeys show imitative behaviour in the first place.[5]
Development
Human infant data using eye-tracking measures suggest that the mirror neuron system develops before 12 months of age, and that this system may help human infants understand other people's actions.[34] A critical question concerns how mirror neurons acquire mirror properties. Two closely related models postulate that mirror neurons are trained through Hebbian[35] or Associative learning[36][37][38] (see Associative Sequence Learning). However, if premotor neurons need to be trained by action in order to acquire mirror properties, it is unclear how newborn babies are able to mimic the facial gestures of another person (imitation of unseen actions), as suggested by the work of Meltzoff and Moore. One possibility is that the sight of tongue protrusion recruits an innate releasing mechanism in neonates. Careful analysis suggests that 'imitation' of this single gesture may account for almost all reports of facial mimicry by new-born infants.[39]
Possible functions
Understanding intentions
Many studies link mirror neurons to understanding goals and intentions. Fogassi et al. (2005)[40] recorded the activity of 41 mirror neurons in the inferior parietal lobe (IPL) of two rhesus macaques. The IPL has long been recognized as an association cortex that integrates sensory information. The monkeys watched an experimenter either grasp an apple and bring it to his mouth or grasp an object and place it in a cup.
- In total, 15 mirror neurons fired vigorously when the monkey observed the "grasp-to-eat" motion, but registered no activity while exposed to the "grasp-to-place" condition.
- For 4 other mirror neurons, the reverse held true: they activated in response to the experimenter eventually placing the apple in the cup but not to eating it.
Only the type of action, and not the kinematic force with which models manipulated objects, determined neuron activity. It was also significant that neurons fired before the monkey observed the human model starting the second motor act (bringing the object to the mouth or placing it in a cup). Therefore, IPL neurons "code the same act (grasping) in a different way according to the final goal of the action in which the act is embedded".[40] They may furnish a neural basis for predicting another individual’s subsequent actions and inferring intention.[40]
Empathy
Stephanie Preston and Frans de Waal,[41] Jean Decety,[42][43] and Vittorio Gallese[44][45] and Christian Keysers[3] have independently argued that the mirror neuron system is involved in empathy. A large number of experiments using functional MRI, electroencephalography (EEG) and magnetoencephalography (MEG) have shown that certain brain regions (in particular the anterior insula, anterior cingulate cortex, and inferior frontal cortex) are active when people experience an emotion (disgust, happiness, pain, etc.) and when they see another person experiencing an emotion.[46][47][48][49][50][51][52] However, these brain regions are not quite the same as the ones which mirror hand actions, and mirror neurons for emotional states or empathy have not yet been described in monkeys.
More recently, Christian Keysers at the Social Brain Lab and colleagues have shown that people who are more empathic according to self-report questionnaires have stronger activations both in the mirror system for hand actions[53] and the mirror system for emotions,[51] providing more direct support for the idea that the mirror system is linked to empathy.
Human self awareness
V.S Ramachandran has speculated that mirror neurons may provide the neurological basis of human self awareness.[54] In an essay written for the Edge Foundation in 2009 Ramachandran gave the following explanation of his theory: “... I also speculated that these neurons can not only help simulate other people's behavior but can be turned "inward"—as it were—to create second-order representations or meta-representations of your own earlier brain processes. This could be the neural basis of introspection, and of the reciprocity of self awareness and other awareness. There is obviously a chicken-or-egg question here as to which evolved first, but... The main point is that the two co-evolved, mutually enriching each other to create the mature representation of self that characterizes modern humans." [55]
Language
In humans, functional MRI studies have reported finding areas homologous to the monkey mirror neuron system in the inferior frontal cortex, close to Broca's area, one of the hypothesized language regions of the brain. This has led to suggestions that human language evolved from a gesture performance/understanding system implemented in mirror neurons. Mirror neurons have been said to have the potential to provide a mechanism for action-understanding, imitation-learning, and the simulation of other people's behaviour.[56] This hypothesis is supported by some cytoarchitectonic homologies between monkey premotor area F5 and human Broca's area.[57] Rates of vocabulary expansion link to the ability of children to vocally mirror non-words and so to acquire the new word pronunciations. Such speech repetition occurs automatically, fast[58] and separately in the brain to speech perception.[59][60] Moreover such vocal imitation can occur without comprehension such as in speech shadowing[61] and echolalia.[62]
Further evidence for this link comes from a recent study in which the brain activity of two participants was measured using fMRI while they were gesturing words to each other using hand gestures with a game of charades – a modality that some have suggested might represent the evolutionary precursor of human language. Analysis of the data using Granger Causality revealed that the mirror-neuron system of the observer indeed reflects the pattern of activity of the activity in the motor system of the sender, supporting the idea that the motor concept associated with the words is indeed transmitted from one brain to another using the mirror system [63]
It must be noticed that the mirror neuron system seems to be inherently inadequate to play any role in syntax, given that this definitory property of human languages which is implemented in hierarchical recursive structure is flattened into linear sequences of phonemes making the recursive structure not accessible to sensory detection[64]
Automatic Imitation
The term is commonly used to refer to cases in which an individual, having observed a body movement, deliberately performs a topographically similar body movement. Automatic Imitation rarely involves overt behavioral execution of matching responses. Automatic imitation effects typically consist of RT, rather than accuracy, differences between compatible and incompatible trials. Research reveals that the existence of automatic imitation, which is a covert form of imitation, is distinct from spatial compatibility. It also indicates that, although automatic imitation subject to input modulation by attentional processes, and output modulation by inhibitory processes, it is mediated by learned, long-term sensorimotor associations that cannot be altered directly by intentional processes. Many researchers believe that automatic imitation is mediated by the mirror neuron system.[65]
Motor Mimicry
In contrast with automatic imitation, motor mimicry is observed in (1) naturalistic social situations and (2) via measures of action frequency within a session rather than measures of speed and/or accuracy within trials. [66]
The integration of research on motor mimicry and automatic imitation could reveal plausible indications that these phenomena depend on the same psychological and neural processes. Preliminary evidence however comes from studies showing that social priming has similar effects on motor mimicry.[67] [68]
Nevertheless, the similarities between automatic imitation, mirror effects, and motor mimicry have led some researchers to propose that automatic imitation is mediated by the mirror neuron system and that it is a tightly controlled laboratory equivalent of the motor mimicry observed in naturalistic social contexts. If true, then automatic imitation can be used as a tool to investigate how the mirror neuron system contributes to cognitive functioning and how motor mimicry promotes prosocial attitudes and behavior.[69][70]
Autism
Some researchers claim there is a link between mirror neuron deficiency and autism. EEG recordings from motor areas are suppressed when someone watches another person move, a signal that may relate to mirror neuron system. This suppression was less in children with autism.[9] Although these findings have been replicated by several groups,[71][72] other studies have not found evidence of a dysfunctional mirror neuron system in autism.[5] In 2008, Oberman et al. published a research paper that presented conflicting EEG evidence. Oberman and Ramachandran found typical mu-suppression for familiar stimuli, but not for unfamiliar stimuli, leading them to conclude that the mirror neuron system of children with ASD was functional, but less sensitive than that of typical children.[73] Based on the conflicting evidence presented by mu-wave suppression experiments, Patricia Churchland has cautioned that mu-wave suppression results cannot be used as a valid index for measuring the performance of mirror neuron systems.[74] Finally, anatomical differences have been found in the mirror neuron related brain areas in adults with autism spectrum disorders, compared to non-autistic adults. All these cortical areas were thinner and the degree of thinning was correlated with autism symptom severity, a correlation nearly restricted to these brain regions.[75] Based on these results, some researchers claim that autism is caused by impairments in the mirror neuron system, leading to disabilities in social skills, imitation, empathy and theory of mind.
Many researchers have pointed out that the "broken mirrors" theory of autism is overly simplistic, and mirror neurons alone cannot explain the deficits found in individuals with autism. First of all, as noted above, none of these studies were direct measures of mirror neuron activity - in other words fMRI activity or EEG rhythm suppression do not unequivocally index mirror neurons. Dinstein and colleagues found normal mirror neuron activity in people with autism using fMRI.[76] In individuals with autism, deficits in intention understanding, action understanding and biological motion perception (the key functions of mirror neurons) are not always found,[77][78] or are task dependent.[79][80] Today, very few people believe an all-or-nothing problem with the mirror system can underlie autism. Instead, "additional research needs to be done, and more caution should be used when reaching out to the media".[81]
Theory of mind
In Philosophy of mind, mirror neurons have become the primary rallying call of simulation theorists concerning our 'theory of mind.' 'Theory of mind' refers to our ability to infer another person's mental state (i.e., beliefs and desires) from experiences or their behaviour. For example, if you see a girl reaching into a jar labeled 'cookies,' you might assume that she wants a cookie and believes that there are cookies in the jar (even if you know the jar is empty).
There are several competing models which attempt to account for our theory of mind; the most notable in relation to mirror neurons is simulation theory. According to simulation theory, theory of mind is available because we subconsciously empathize with the person we're observing and, accounting for relevant differences, imagine what we would desire and believe in that scenario.[82][83] Mirror neurons have been interpreted as the mechanism by which we simulate others in order to better understand them, and therefore their discovery has been taken by some as a validation of simulation theory (which appeared a decade before the discovery of mirror neurons).[84] More recently, Theory of Mind and Simulation have been seen as complementary systems, with different developmental time courses.[85][86][87]
Gender differences
The issue of gender differences in empathy is quite controversial and subject to social desirability and stereotypes. However, a series of recent studies conducted by Yawei Cheng, using a variety of neurophysiological measures, including MEG,[88] spinal reflex excitability,[89] electroencephalography,[90][91] have documented the presence of a gender difference in the human mirror neuron system, with female participants exhibiting stronger motor resonance than male participants.
Criticism
Although many in the scientific community have been excited about the discovery of mirror neurons, there are some researchers who express skepticism in regards to the claims that mirror neurons can explain empathy, theory of mind, etc. Greg Hickok, a cognitive neuroscientist at UC Irvine, has claimed that "there is little or no evidence to support the 'mirror neuron = action understanding' hypothesis".[92]
See also
- Associative Sequence Learning
- Common coding theory
- Emotional contagion
- Empathy
- Motor cognition
- Motor theory of speech perception
- On Intelligence
- Positron emission tomography
- Simulation theory of empathy
- Speech repetition
- Spindle neuron
References
- ^ a b c d Rizzolatti, Giacomo; Craighero, Laila (2004). "The mirror-neuron system". Annual Review of Neuroscience 27: 169–192. doi:10.1146/annurev.neuro.27.070203.144230. PMID 15217330. http://www.kuleuven.be/mirrorneuronsystem/readinglist/Rizzolatti%20&%20Craighero%202004%20-%20The%20MNS%20-%20ARN.pdf
- ^ Keysers, Christian (2010). "Mirror Neurons". Current Biology 19 (21): R971–973. doi:10.1016/j.cub.2009.08.026. PMID 19922849. http://www.bcn-nic.nl/txt/people/publications/2009_Keysers_CurrentBiology.pdf.
- ^ a b c d Keysers, Christian (2011-06-23). The Empathic Brain. Kindle. http://www.facebook.com/theempathicbrain.
- ^ V.S. Ramachandran, "Mirror Neurons and imitation learning as the driving force behind "the great leap forward" in human evolution". Edge Foundation. http://www.edge.org/3rd_culture/ramachandran/ramachandran_p1.html. Retrieved 2006-11-16.
- ^ a b c d e Dinstein I, Thomas C, Behrmann M, Heeger DJ (2008). "A mirror up to nature". Curr Biol 18 (1): R13–8. doi:10.1016/j.cub.2007.11.004. PMC 2517574. PMID 18177704. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2517574.
- ^ Christian Keysers and Valeria Gazzola, Progress in Brain Research, 2006, [1]
- ^ Michael Arbib, The Mirror System Hypothesis. Linking Language to Theory of Mind, 2005, retrieved 2006-02-17
- ^ Théoret, Hugo; Pascual-Leone, Alvaro (2002). "Language Acquisition: Do as You Hear". Current Biology 12 (21): R736–7. doi:10.1016/S0960-9822(02)01251-4. PMID 12419204.
- ^ a b Oberman LM, Hubbard EM, McCleery JP, Altschuler EL, Ramachandran VS, Pineda JA., EEG evidence for mirror neuron dysfunction in autism spectral disorders, Brain Res Cogn Brain Res.; 24(2):190-8, 2005-06
- ^ Dapretto, M; Davies, MS; Pfeifer, JH; Scott, AA; Sigman, M; Bookheimer, SY; Iacoboni, M (2006). "Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders". Nature neuroscience 9 (1): 28–30. doi:10.1038/nn1611. PMID 16327784.
- ^ Di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G. (1992). Understanding motor events: a neurophysiological study. Experimental Brain Research, 91, 176-180.
- ^ Giacomo Rizzolatti et al. (1996) Premotor cortex and the recognition of motor actions, Cognitive Brain Research 3 131-141
- ^ Rizzolatti G, Fabbri-Destro M (2010). "Mirror neurons: from discovery to autism". Exp Brain Res 200 (3–4): 223–37. doi:10.1007/s00221-009-2002-3. PMID 19760408.
- ^ Gallese, V.; Fadiga, L.; Fogassi, L.; Rizzolatti, Giacomo (1996). "Action recognition in the premotor cortex". Brain 119 (2): 593–609. doi:10.1093/brain/119.2.593. PMID 8800951. http://brain.oxfordjournals.org/cgi/content/abstract/119/2/593
- ^ Ferrari et al. (2003) "Mirror neurons responding to the observation of ingestive and communicative mouth actions in the ventral premotor cortex. European Journal of Neuroscience, 17 (8), 1703-1714
- ^ Kohler et al., Science, 2002 [2]
- ^ Gazzola et al., Current Biology, 2006 [3]
- ^ Gallese et al., Action recognition in the premotor cortex, Brain, 1996
- ^ Fogassi et al., Parietal Lobe: From Action Organization to Intention Understanding, Science, 2005
- ^ a b c Gazzola, V.; Keysers, C. (2009). "The observation and execution of actions share motor and somatosensory voxels in all tested subjects: single-subject analyses of unsmoothed fMRI data". Cereb Cortex 19 (6): 1239–1255. doi:10.1093/cercor/bhn181. PMC 2677653. PMID 19020203. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2677653
- ^ Keysers, Christian; Kaas, John; Gazzola, Valeria (2010). "Somatosensation in Social Cognition". Nature Reviews Neuroscience 11 (6): 417–28. doi:10.1038/nrn2833. PMID 20445542. http://www.bcn-nic.nl/txt/people/publications/2010_NRN2833_Keysers_Kaas_Gazzola.pdf.
- ^ Ferrari, P. F.; Visalberghi, E.; Paukner, A.; Fogassi, L.; Ruggiero, A.; Suomi, SJ (2006). et al.. "Neonatal Imitation in Rhesus Macaques". PLoS Biology 4 (9): e302. doi:10.1371/journal.pbio.0040302. PMC 1560174. PMID 16953662. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1560174
- ^ Ferrari P.F., Bonini L., Fogassi L. (2009) From monkey mirror neurons to mirror-related behaviours: possible direct and indirect pathways. Philosophical Transactions of the Royal Society, B. .364, 2311-2323
- ^ Rizzolatti, Giacomo; Arbib, Michael A. (1998). "Language within our grasp". Trends in Neurosciences 21 (5): 188–194. doi:10.1016/S0166-2236(98)01260-0. PMID 9610880
- ^ Iacoboni, Marco; Woods, Roger P.; Brass, Marcel; Bekkering, Harold; Mazziotta, John C.; Rizzolatti, Giacomo (1999). "Cortical Mechanisms of Human Imitation". Science 286 (5449): 2526–2528. doi:10.1126/science.286.5449.2526. PMID 10617472
- ^ Saygin AP, Wilson SM, Dronkers NF, Bates E (2004). "Action comprehension in aphasia: linguistic and non-linguistic deficits and their lesion correlates". Neuropsychologia 42 (13): 1788-804. doi:10.1016/j.neuropsychologia.2004.04.016. PMID 15351628.
- ^ a b Tranel, D; Kemmerer, D; Adolphs, R; Damasio, H; Damasio, AR (2003 May 1). "Neural correlates of conceptual knowledge for actions". Cognitive Neuropsychology 20 (3): 409–32. doi:10.1080/02643290244000248. PMID 20957578.
- ^ a b Saygin, A. P. (2007). "Superior temporal and premotor brain areas necessary for biological motion perception". Brain : a journal of neurology 130 (Pt 9): 2452–2461. doi:10.1093/brain/awm162. PMID 17660183.
- ^ a b Pobric G; Hamilton AF (2006 Mar 7). "Action understanding requires the left inferior frontal cortex". Current Biology 16 (5): 524–9. doi:10.1016/j.cub.2006.01.033. PMID 16527749.
- ^ Candidi M; Urgesi C; Ionta S; Aglioti SM (2008). "Virtual lesion of ventral premotor cortex impairs visual perception of biomechanically possible but not impossible actions". Social Neuroscience 3 (3-4): 388–400. doi:10.1080/17470910701676269. PMID 18979387.
- ^ Keysers, Christian; Gazzola, Valeria (2010). "Social Neuroscience: Mirror Neurons recorded in Humans". Current Biology 20 (8): R353–354. doi:10.1016/j.cub.2010.03.013. PMID 21749952. http://www.bcn-nic.nl/txt/people/publications/2010_KeysersGazzolaMirrorNeuronsRecordedInHumans.pdf.
- ^ a b Pascolo PB, Ragogna R, Rossi R, (2009). "The Mirror-Neuron System Paradigm and its consistency". Gait Posture 30 (Suppl. 1): 65. doi:10.1016/j.gaitpost.2009.07.064.
- ^ a b Hickok G (2009). "Eight problems for the mirror neuron theory of action understanding in monkeys and humans". J Cogn Neurosci 21 (7): 1229–1243. doi:10.1162/jocn.2009.21189. PMC 2773693. PMID 19199415. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2773693.
- ^ Terje Falck-Ytter, Gustaf Gredebäck & Claes von Hofsten (2006), Infants predict other people's action goals[4], Nature Neuroscience 9 (2006)
- ^ Keysers & Perrett, Trends in Cognitive Sciences 8 (2004)
- ^ Heyes C. M. (2001). "Causes and consequences of imitation". Trends in Cognitive Sciences 5 (6): 253–261. doi:10.1016/S1364-6613(00)01661-2. PMID 11390296.
- ^ Brass, M., & Heyes, C. Trends in Cognitive Sciences 9 (2005)
- ^ Heyes, C. M. (2010) Where do mirror neurons come from? Neuroscience and Biobehavioural Reviews, 34, 575-583
- ^ Anisfeld, M. (1996). Only tongue protruding modeling is matched by neonates. Developmental Review, 16(2), 149-161
- ^ a b c Fogassi Leonardo, Pier , Ferrari Francesco, Gesierich Benno, Rozzi Stefano, Chersi Fabian, Rizzolatti Giacomo (2005). "Parietal lobe: from action organization to intention understanding". Science 308 (5722): 662–667. doi:10.1126/science.1106138. PMID 15860620.
- ^ Preston, S. D., & de Waal, F.B.M. (2002) Empathy: Its ultimate and proximate bases. Behavioral and Brain Sciences, 25, 1-72.
- ^ Decety, J. (2002). Naturaliser l’empathie [Empathy naturalized]. L’Encéphale, 28, 9-20.
- ^ Decety, J., & Jackson, P.L. (2004). The functional architecture of human empathy. Behavioral and Cognitive Neuroscience Reviews, 3, 71-100.
- ^ Gallese V., Goldman A.I. (1998). "Mirror neurons and the simulation theory". Trends in Cognitive Sciences 2 (12): 493–501. doi:10.1016/S1364-6613(98)01262-5. PMID 21227300.
- ^ Gallese V (2001). "The "Shared Manifold" hypothesis: from mirror neurons to empathy". Journal of Consciousness Studies 8: 33–50.
- ^ Botvinick M., Jha A.P., Bylsma L.M., Fabian S.A., Solomon P.E., Prkachin K.M. (2005). "Viewing facial expressions of pain engages cortical areas involved in the direct experience of pain". NeuroImage 25 (1): 312–319. doi:10.1016/j.neuroimage.2004.11.043. PMID 15734365.
- ^ Cheng Y., Yang C.Y., Lin C.P., Lee P.R., Decety J. (2008). "The perception of pain in others suppresses somatosensory oscillations: a magnetoencephalography study". NeuroImage 40 (4): 1833–1840. doi:10.1016/j.neuroimage.2008.01.064. PMID 18353686.
- ^ Morrison, I., Lloyd, D., di Pellegrino, G., & Roberts, N. (2004). Vicarious responses to pain in anterior cingulate cortex: is empathy a multisensory issue? Cognitive and Affective Behavioral Neuroscience, 4, 270-278.
- ^ Wicker et al., Neuron, 2003 [5]
- ^ Singer et al., Science, 2004 [6]
- ^ a b Jabbi, Swart and Keysers, NeuroImage, 2006 [7]
- ^ Lamm C., Batson C.D., Decety J. (2007). "The neural substrate of human empathy: effects of perspective-taking and cognitive appraisal". Journal of Cognitive Neuroscience 19 (1): 42–58. doi:10.1162/jocn.2007.19.1.42. PMID 17214562.
- ^ Gazzola, Aziz-Zadeh and Keysers, Current Biology, 2006 [8]
- ^ Oberman, L.; Ramachandran, V.S. (2009). "Reflections on the Mirror Neuron System: Their Evolutionary Functions Beyond Motor Representation". In Pineda, J.A.. Mirror Neuron Systems: The Role of Mirroring Processes in Social Cognition. Humana Press. pp. 39–62. ISBN 978-1-934115-34-3.
- ^ Ramachandran, V.S. (January 1, 2009). "Self Awareness: The Last Frontier, Edge Foundation web essay". http://www.edge.org/3rd_culture/rama08/rama08_index.html. Retrieved July 26, 2011.
- ^ Skoyles, John R., Gesture, Language Origins, and Right Handedness, Psycholoqy: 11,#24, 2000
- ^ Petrides, Michael, Cadoret, Genevieve, Mackey, Scott (2005). Orofacial somatomotor responses in the macaque monkey homologue of Broca's area, Nature: 435,#1235
- ^ Porter RJ. Lubker JF. (1980). Rapid reproduction of vowel-vowel sequences, Evidence for a fast and direct acoustic-motoric linkage in speech. Journal of Speech and Hearing Research, 23, 593-602. PubMed
- ^ McCarthy R. Warrington EK. (1984). A two-route model of speech production, Evidence from aphasia. Brain, 107, 463-485. PubMed
- ^ McCarthy RA, Warrington EK. (2001). Repeating without semantics: surface dysphasia? Neurocase.;7:77-87. PubMed
- ^ Marslen-Wilson W. (1973). Linguistic structure and speech shadowing at very short latencies. Nature, 244, 522-523. PubMed
- ^ Fay WH. Coleman RO. (1977). A human sound transducer/reproducer: temporal capabilities of a profoundly echolalic child. Brain and Language, 4, 396-402. PubMed
- ^ Schippers, MB; Roebroeck, A; Renken, R; Nanetti, L; Keysers, C (2010). "Mapping the Information flow from one brain to another during gestural communication". Proc Natl Acad Sci U S A. 107 (20): 9388–93. doi:10.1073/pnas.1001791107. PMC 2889063. PMID 20439736. http://www.bcn-nic.nl/txt/people/publications/2010_SchippersKeysers_PNAS.pdf.
- ^ Moro, Andrea (2008). The Boundaries of Babel. The Brain and the Enigma of Impossible Languages. MIT Press. pp. 257. ISBN 13: 978-0-262-13498-9.. http://mitpress.mit.edu/catalog/item/default.asp?ttype=2&tid=11488.
- ^ Longo, M. R; Kosobud, A., & Bertenthal, B. I. (2008). "Human Perception and Performance". Journal of Experimental Psychology.
- ^ Chartrand, T. L; Bargh, J. A (1999). Journal of Personality and Social Psychology.
- ^ Lakin, J. L; Chartrand, T. L (2003). Psychological Science.
- ^ van Baaren, R. B; Maddux, W. W., Chartrand, T. L., De Bouter, C., & van Knippenberg, A (2003). Journal of Personality and Social Psychology.
- ^ Heyes, Cecilia (2011). Psychological Bulletin 137.
- ^ Paukner A., Visalberghi E., Suomi S.J., Ferrari P.F. (2009). Capuchin monkeys display affiliation towards humans who imitate them. Science, 325, 880-883
- ^ Dapretto M, Davies MS, Pfeifer JH et al. (January 2006). "Understanding emotions in others: mirror neuron dysfunction in children with autism spectrum disorders". Nat. Neurosci. 9 (1): 28–30. doi:10.1038/nn1611. PMID 16327784.
- ^ Perkins T, Stokes M, McGillivray J, Bittar R. (2010). "Mirror neuron dysfunction in autism spectrum disorders". J. Clin. Neurosci. 17 (10): 1239–1243. doi:10.1016/j.jocn.2010.01.026. PMID 20598548.
- ^ Lindsay M. Oberman, Vilayanur S. Ramachandran, Jaime A. Pineda (2008). "Modulation of mu suppression in children with autism spectrum disorders in response to familiar or unfamiliar stimuli: The mirror neuron hypothesis". Neuropsychologia 46 (5): 1558–1565. doi:10.1016/j.neuropsychologia.2008.01.010. PMID 18304590. http://tmslab.org/wp-content/files/familiaritypublication.pdf.
- ^ Churcland, P.S. (2011). "6". Braintrust. Princeton University Press. p. 156.
- ^ Hadjikhani and others; Joseph, RM; Snyder, J; Tager-Flusberg, H (2006). "Anatomical Differences in the Mirror Neuron System and Social Cognition Network in Autism". Cerebral Cortex 16 (9): 1276–82. doi:10.1093/cercor/bhj069. PMID 16306324.
- ^ http://www.newscientist.com/article/dn18837-mirror-neurons-seen-behaving-normally-in-autism.html
- ^ Hamilton, AF (2009 Aug). "Goals, intentions and mental states: challenges for theories of autism". Journal of Child Psychology and Psychiatry 50 (8): 881–92. doi:10.1111/j.1469-7610.2009.02098.x. PMID 19508497.
- ^ Murphy, P; Brady, N; Fitzgerald, M; Troje, NF (2009). "No evidence for impaired perception of biological motion in adults with autistic spectrum disorders". Neuropsychologia 47 (14): 3225–35. doi:10.1016/j.neuropsychologia.2009.07.026. PMID 19666038.
- ^ Saygin, A.; Cook, J.; Blakemore, S. (2010 Oct 18). Baker, Chris I. ed. "Unaffected perceptual thresholds for biological and non-biological form-from-motion perception in autism spectrum conditions". PLoS ONE 5 (10): e13491. Bibcode 2010PLoSO...513491S. doi:10.1371/journal.pone.0013491. PMC 2956672. PMID 20976151. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2956672.
- ^ Cook, Jennifer; Saygin, Ayse Pinar; Swain, Rachel; Blakemore, Sarah-Jayne (2009). "Reduced sensitivity to minimum-jerk biological motion in autism spectrum conditions". Neuropsychologia 47 (14): 3275–3278. doi:10.1016/j.neuropsychologia.2009.07.010. PMC 2779370. PMID 19632248. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2779370.
- ^ Fan YT; Decety J; Yang CY; Liu JL; Cheng Y (2010 Sep). "Unbroken mirror neurons in autism spectrum disorders". Journal of Child Psychology and Psychiatry 51 (9): 981–8. doi:10.1111/j.1469-7610.2010.02269.x. PMID 20524939.
- ^ Gordon, R. (1986). Folk psychology as simulation. Mind and Language 1: 158-171
- ^ Goldman, A. (1989). Interpretation psychologized. Mind and Language 4: 161–185
- ^ Gallese V., Goldman A. (1998). "Mirror neurons and the simulation theory of mindreading". Trends in Cognitive Sciences 2 (12): 493–501. doi:10.1016/S1364-6613(98)01262-5. PMID 21227300.
- ^ Meltzoff, A.N., & Decety, J. (2003). What imitation tells us about social cognition: A rapprochement between developmental psychology and cognitive neuroscience. The Philosophical Transactions of the Royal Society, London, 358, 491-500.
- ^ Sommerville, J. A., & Decety, J. (2006). Weaving the fabric of social interaction: Articulating developmental psychology and cognitive neuroscience in the domain of motor cognition. Psychonomic Bulletin & Review, 13, 179-200.
- ^ Keysers C and Gazzola V(2007) Integrating simulation and theory of mind: from self to social cognition. Trends in Cognitive Sciences 11(5):194-6 [9]
- ^ Cheng Y., Tzeng O.J., Decety J., Hsieh J.C. (2006). "Gender differences in the human mirror system: a magnetoencephalography study". NeuroReport 17 (11): 1115–1119. doi:10.1097/01.wnr.0000223393.59328.21. PMID 16837838.
- ^ Cheng Y., Decety J., Hsieh J.C., Hung D., Tzeng O.J. (2007). "Gender differences in spinal excitability during observation of bipedal locomotion". NeuroReport 18 (9): 887–890. doi:10.1097/WNR.0b013e3280ebb486. PMID 17515795.
- ^ Cheng, Y., Decety, J., Yang, C.Y., Lee, S., & Chen, G. (2008). Gender differences in the Mu rhythm during empathy for pain: An electroencephalographic study. Brain Research, in press.
- ^ Cheng Y., Lee P., Yang C.Y., Lin C.P., Decety J. (2008). "Gender differences in the mu rhythm of the human mirror-neuron system". PLoS ONE 3 (5): e2113. doi:10.1371/journal.pone.0002113. PMC 2361218. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2361218.
- ^ Greg Hickok (2009-02-10). "Eight problems for the mirror neuron theory of action understanding". Talking Brains blog. http://talkingbrains.blogspot.com/2009/02/eight-problems-for-mirror-neuron-theory.html. Retrieved 2010-08-29.
Further reading
- Keysers, C. (2011) The Empathic Brain. Amazon [10]
- Iacoboni M, Mazziotta JC (2007). "Mirror neuron system: basic findings and clinical applications". Ann Neurol 62 (3): 213–8. doi:10.1002/ana.21198. PMID 17721988.
- Keysers, C., & Gazzola, V. (2006), Towards a unifying neural theory of social cognition, Progress in Brain Research.[11]
- Morsella, E., Bargh, J.A., & Gollwitzer, P.M. (Eds.) (2009). Oxford Handbook of Human Action. New York: Oxford University Press.
- Preston, S. D., & de Waal, F.B.M. (2002). Empathy: Its ultimate and proximate bases. Behavioral and Brain Sciences, 25, 1-72.
- Rizzolatti G, Fabbri-Destro M, Cattaneo L (2009). "Mirror neurons and their clinical relevance". Nat Clin Pract Neurol 5 (1): 24–34. doi:10.1038/ncpneuro0990. PMID 19129788. http://www.nature.com/ncpneuro/journal/v5/n1/full/ncpneuro0990.html.
- Rizzolatti, G, Sinigaglia, C. (2008). Mirrors in the Brain. How We Share our Actions and Emotions. Oxford University Press.
External links
- NOVA scienceNOW: Mirror Neurons (including a 14 minute broadcast segment)
- You remind me of me in the New York Times.
- 7 Minute Video on TED.com :: Neuroscientist Vilayanur Ramachandran outlines the functions of mirror neurons
- Mirror Neuron Forum, Perspectives on Psychological Science, September, 2011
Categories:- CNS neurons
- Cognitive neuroscience
- Behavioral neuroscience
- Motor control
- Cognitive science
- Neuroscience


Wikimedia Foundation. 2010.