- Megalodon
-
For other uses, see Megalodon (disambiguation).
Megalodon
Temporal range: Late Oligocene-Early Pleistocene 28–1.5 Ma
Model of the jaws of the megalodon at the American Museum of Natural History. Scientific classification Kingdom: Animalia Phylum: Chordata Subphylum: Vertebrata Class: Chondrichthyes Subclass: Elasmobranchii Order: Lamniformes Family: Disputed; Lamnidae or †Otodontidae Genus: Disputed; Carcharodon or †Carcharocles Species: †C. megalodon Binomial name Disputed; Carcharodon megalodon or Carcharocles megalodon
For Carcharodon megalodon, Agassiz, 1843Synonyms - Procarcharodon megalodon Casier, 1960
- Megaselachus megalodon Glikman, 1964
The megalodon (
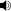 /ˈmɛɡələdɒn/ meg-ə-lə-don; meaning "big tooth", from Greek μέγας (mega, "big") and ὀδούς (odon, "tooth")) is an extinct species of shark that lived roughly from 28 to 1.5 million years ago, during the Cenozoic Era (late Oligocene to early Pleistocene).
/ˈmɛɡələdɒn/ meg-ə-lə-don; meaning "big tooth", from Greek μέγας (mega, "big") and ὀδούς (odon, "tooth")) is an extinct species of shark that lived roughly from 28 to 1.5 million years ago, during the Cenozoic Era (late Oligocene to early Pleistocene).The taxonomic assignment of C. megalodon has been debated for nearly a century, and is still under dispute with two major interpretations; Carcharodon megalodon (under family Lamnidae) or Carcharocles megalodon (under family Otodontidae).[1] Consequently, the scientific name of this species has been commonly abbreviated to C. megalodon in literature.
C. megalodon is regarded as one of the largest and most powerful predators in vertebrate history.[2] C. megalodon likely had a profound impact on structuring of the marine communities. Fossil remains indicate that this giant shark reached a total length (TL) of more than 16 metres (52 ft),[1] and also affirm that it had a cosmopolitan distribution.[1] Scientists suggest that C. megalodon looked like a stockier version of the great white shark, Carcharodon carcharias, in life.
Contents
Discovery
Glossopetrae
 The depiction of a shark's head by Nicolaus Steno in his work, The Head of a Shark Dissected.
The depiction of a shark's head by Nicolaus Steno in his work, The Head of a Shark Dissected.
According to Renaissance accounts, gigantic, triangular fossil teeth often found embedded in rocky formations were once believed to be petrified tongues, or glossopetrae, of the dragons and snakes. This interpretation was corrected in 1667 by a Danish naturalist, Nicolaus Steno, who recognized them as ancient shark teeth (and famously produced a depiction of a shark's head bearing such teeth).[3] He mentioned his findings in a book, The Head of a Shark Dissected, which also contained an illustration of a C. megalodon tooth, previously considered to be a tongue stone.[4]
Identification
A Swiss naturalist, Louis Agassiz, gave this shark its scientific name, Carcharodon megalodon, in 1835,[5] in his research work Recherches sur les poissons fossiles[6] (Research on fossil fish), which he completed in 1843. The teeth of the C. megalodon are morphologically similar to the teeth of the great white shark. On the basis of this observation, Agassiz assigned the genus Carcharodon to the megalodon.[5] While the scientific name is C. megalodon, it is often informally dubbed the megatooth shark[7] or giant white shark[8] or even monster shark.[9]
Fossils
C. megalodon is represented in the fossil record primarily by teeth and vertebral centra.[7] As with all other sharks, C. megalodon's skeleton was formed of cartilage rather than bone; this results in mostly poorly preserved fossil specimens.[10]
Fossil teeth
 Megalodon tooth with slant height (diagonal length) of over 170 mm.
Megalodon tooth with slant height (diagonal length) of over 170 mm.The most common fossils of C. megalodon are its teeth. The diagnostic characters of C. megalodon teeth include: triangular shape,[1] robust structure,[7] large size,[1] fine serrations,[1] and visible v-shaped neck.[1] The teeth of C. megalodon can measure over 180 millimetres (7.1 in) in slant height or diagonal length, and are the largest in size of any known shark species.[11]
Fossil vertebrae
Fossil vertebrae of C. megalodon have also been occasionally found.[4] The most notable example is a partially preserved but associated vertebral column of a single C. megalodon specimen, which was excavated from Antwerp basin, Belgium by M. Leriche in 1926. This specimen comprises 150 vertebral centra, with the largest centra being 155 mm in diameter.[7] However, scientists have claimed that considerably larger vertebral centra can be expected from C. megalodon.[7] Interestingly, a partially preserved but associated vertebral column of another C. megalodon specimen was excavated from Gram clay, Denmark by Bendix-Almgeen in 1983. This specimen comprises 20 vertebral centra, with the largest centra being around 230 mm in diameter.[12]
Fossil distribution and age
The fossils of C. megalodon have been excavated from many parts of the world, including Europe,[4] North America,[7] South America,[4][7] Puerto Rico,[13] Cuba,[14] Jamaica,[15] Australia,[16] New Zealand,[11] Japan,[4][7] Africa,[4][7] Malta,[11] Grenadines,[17] and India.[4] C. megalodon teeth have also been excavated from regions far away from continental lands (e.g., the Mariana Trench in the Pacific).[11]
The earliest remains of C. megalodon have been reported from late Oligocene strata, circa 28 million years old.[18][19] Although fossils of C. megalodon are predominantly absent in strata extending beyond the Tertiary boundary,[7] they have been reported from subsequent Pleistocene strata.[20] It is believed that C. megalodon became extinct in the Pleistocene, probably about 1.5 million years ago.[10]
Anatomy
Among extant species, the great white shark is regarded as the best analogue to C. megalodon.[1] The lack of well preserved fossil skeletons of C. megalodon have forced scientists to rely on the morphology of the great white shark for the basis of its reconstruction and size estimation.[7]
Size estimation
Due to fragmentary remains, estimating the size of C. megalodon has been challenging.[11] However, the scientific community acknowledges that C. megalodon was larger than the whale shark, Rhincodon typus. Scientists have focused their research upon two aspects of size: (1) total length (TL), and (2) body mass (BM).
Total length estimates
The first attempt to reconstruct the jaw of this shark was made by Professor Bashford Dean in 1909. From the dimensions of this jaw reconstruction, it was hypothesized that C. megalodon could have approached 30 metres (98 ft) in total length (TL), but in light of new fossil discoveries and advances in vertebrate sciences this jaw reconstruction is now considered to be inaccurate.[21] Major reasons cited for this inaccuracy are (1) relatively poor knowledge of C. megalodon's dentition in Dean's time, and (2) inaccurate muscle structures.[21] Experts suggest that a rectified version of C. megalodon's jaw model by Bashford Dean would be about seventy percent (70%) of its original size and would lead to a shark size consistent with modern findings.[21] To resolve such errors, scientists, aided by new fossil discoveries of C. megalodon and improved knowledge of its closest living analogue's anatomy, introduced more quantitative methods for estimating its size based on the statistical relationships between the tooth sizes and body lengths in the great white shark.[7][21] Some methods are mentioned below.
Method proposed by John E. Randall
In 1973, an ichthyologist from Hawaii, John E. Randall, introduced a method to estimate the total length of the great white shark.[22] The proposed method is depicted in the form of a plotted graph, which demonstrates a relationship between the enamel height (the vertical distance of the blade from the base of the enamel portion of the tooth to its tip) of the largest tooth in the upper jaw of the great white shark and its total length.[21][22] Randall extrapolated this method to estimate the total length of C. megalodon as well.[22] Randall cited two C. megalodon teeth in his work: (1) specimen number 10356 in American Museum of Natural History and (2) specimen number 25730 in United States National Museum, which had enamel heights of 115 mm and 117.5 mm respectively.[22] These teeth yielded a corresponding total length of about 13 metres (43 ft).[21][22] In 1991, two shark experts, Richard Ellis and John E. McCosker, pointed out a flaw in Randall's method.[7] According to them, sharks' tooth enamel height does not necessarily increase in proportion to the animal's total length. This observation led to proposals for new, more accurate methods to determine the size of the great white shark and similar sharks.[7]
Method proposed by Gottfried et al.
In 1996 three scientists – Michael D. Gottfried, Leonard J. V. Compagno and S. Curtis Bowman – after thorough research and scrutiny of 73 great white shark specimens, proposed a conservative linear relationship between the maximum height of the largest upper anterior tooth (UA) and the total length (TL) in the great white shark.[7][19] The proposed relationship is: total length (TL) in meters = − (0.22) + (0.096) × [UA maximum height (mm)].[7][19] Gottfried and colleagues (1996) also extrapolated this method to estimate the total length (TL) of C. megalodon. The biggest C. megalodon tooth in the possession of this team was an upper second anterior specimen, whose maximum height was 168 millimetres (6.6 in). This tooth had been discovered by Compagno in 1993, and it yielded an estimated total length (TL) of 15.9 metres (52 ft).[7] However, rumors of larger C. megalodon teeth persisted at that time.[7] The maximum tooth height for this method is measured as a vertical line from the tip of the crown to the bottom of the lobes of the root, parallel to the long axis of the tooth.[7] In layman's terms, the maximum height of the tooth is its slant height.[23]
Method proposed by Clifford Jeremiah
In 2002, shark researcher Dr. Clifford Jeremiah proposed a method to estimate the total length (TL) of great white shark and similar sharks including C. megalodon.[11] Shark researcher David Ward has asserted that this method is based on a sound principle that works well with most large sharks.[11] The proposed method is: for every centimeter of root width of an upper anterior tooth, there is approximately 4.5 feet of the shark. Jeremiah pointed out that the jaw perimeter of a shark is directly proportional to its total length (TL), with the width of the roots of the largest teeth being a proxy for estimating jaw perimeter.[11] The largest tooth in the possession of Jeremiah had a root width of about 12 centimetres (4.7 in), which yielded 15.5 metres (51 ft) total length (TL).[11]
Method proposed by Kenshu Shimada
In 2002, paleontologist Dr. Kenshu Shimada of DePaul University proposed a linear relationship between tooth crown height (CH) and total length (TL) in great white sharks after conducting anatomical analysis of several specimens.[24] In statistical terms, this relationship is expressed as: total length (TL) in cm = a + bx, where a is a constant, b is the slope of the line, and x is the crown height (CH) of tooth in mm. This proposed relationship provides the capability to predict total length (TL) on the basis of crown height (CH) of any tooth in great white shark.[1][24] The crown height (CH) of the tooth is measured as maximum vertical enameloid height on the labial side for this method.[24] Shimada pointed out that previously proposed methods were based on weaker evaluation of dental homology.[24] Shimada pointed out that the growth rate between the crown and root is not isometric, and this factor has been taken into consideration in his proposed model.[24] Furthermore, this proposed relationship can also be used to predict the total length (TL) of sharks that are morphologically similar to the great white shark (e.g., C. megalodon).[1][24] Using this model, the upper anterior tooth (with maximum height of 168 millimetres (6.6 in)) possessed by Gottfried and colleagues (1996) corresponded to a total length (TL) of 15.1 metres (50 ft).[24] In 2010, several shark researchers – Catalina Pimiento, Dana J. Ehret, Bruce J. MacFadden, and Gordon Hubbell — estimated the total length (TL) of C. megalodon on the basis of Shimada's method.[1] Among the specimens found in the Gatun Formation of Panama, specimen number 237956 yielded a total length (TL) of 16.8 metres (55 ft).[1]
Consensus
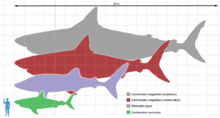 Megalodon (gray and red) with the whale shark (violet), great white shark (green), and a human (blue) for scale. Note: The maximum size attained by C. megalodon is indicated by the 20 m scale.
Megalodon (gray and red) with the whale shark (violet), great white shark (green), and a human (blue) for scale. Note: The maximum size attained by C. megalodon is indicated by the 20 m scale.In the 1990s, marine biologists (e.g., Patrick J. Schembri, and Staphon Papson) opined that C. megalodon may have approached a maximum of around 24 to 25 metres (79 to 82 ft) in total length (TL).[25][26] The early total length (TL) estimation of C. megalodon is perhaps not far-fetched. However, Gottfried and colleagues (1996) proposed that C. megalodon could likely approach a maximum of 20.3 metres (67 ft) in total length (TL).[2][7][27] Currently most experts agree this giant shark reached a total length (TL) of more than 16 metres (52 ft).[1][2][11][27]
Largest known specimens
Dr. Gordon Hubbell from Gainesville in USA, possesses an upper anterior C. megalodon tooth whose maximum height is 184.15 millimetres (7.250 in).[11] In addition, a C. megalodon jaw reconstruction contains a tooth whose maximum height is reportedly 193.675 millimetres (7.6250 in).[28] This jaw reconstruction was developed by deceased fossil hunter Vito Bertucci,[28] who was known as "Megalodon Man".[11][28]
Body mass estimates
Gottfried and colleagues (1996) also introduced a method to determine the body mass of the great white shark after studying the length–mass relationship data of 175 specimens at various growth stages and extrapolated it to estimate the body mass of C. megalodon. The proposed method is: mass in kilogram = 3.29E−06[TL in (meters)3.174].[7] According to this model, a 15.9 metres (52 ft) long C. megalodon would have a body mass of about 47 metric tons (52 short tons),[2][7] a 17 metres (56 ft) long C. megalodon would have a body mass of about 59 metric tons (65 short tons),[7] and a 20.3 metres (67 ft) long C. megalodon would have a body mass of 103 metric tons (114 short tons).[2][7] Consequently, C. megalodon is regarded as the largest shark ever to have lived,[1] and is among the largest fish known to have existed.[7]
Dentition and jaw mechanics
 Reconstruction showing the position of the replacement teeth.
Reconstruction showing the position of the replacement teeth.A team of Japanese scientists, T. Uyeno, O. Sakamoto, and H. Sekine, discovered and excavated partial remains of a C. megalodon, with nearly complete associated set of its teeth, from Saitama, Japan in 1989.[4] Another nearly complete associated C. megalodon dentition was excavated from Yorktown Formations of Lee Creek, North Carolina in USA and served as the basis of a jaw reconstruction of C. megalodon in the American Museum of Natural History in NYC.[7] These associated tooth sets solved the mystery of how many teeth would be in the jaws of the C. megalodon in each row in life. Hence, highly accurate jaw reconstructions were now possible. More associated dentitions of C. megalodon have also been found in later years. Based upon these discoveries, two scientists, S. Applegate and L. Espinosa, published an artificial dental formula (representation of dentition of an animal with respect to types of teeth and their arrangement within the animal's jaw) for C. megalodon in 1996.[4][7] Most accurate modern C. megalodon jaw reconstructions are based on this dental formula.
The dental formula of C. megalodon is:
 .[4]
.[4]As evident from the dental formula, C. megalodon had four kinds of teeth in its jaws.[4]
- Anterior - A
- Intermediate - I (In the case of C. megalodon, this tooth technically appears to be an upper anterior and is termed as "A3" because it is fairly symmetrical and does not points mesially (side of the tooth toward the midline of the jaws where left and right jaws meet), but this tooth is still designated as an intermediate tooth.[5] However, in the case of the great white shark, the intermediate tooth does point mesially. This point has often been raised in the Carcharodon vs. Carcharocles debate regarding the megalodon and favors the case of Carcharocles proponents.)
- Lateral - L
- Posterior - P
 Reconstructed jaws on display at the National Aquarium in Baltimore.
Reconstructed jaws on display at the National Aquarium in Baltimore.C. megalodon had a very robust dentition,[7] and it had a total of about 276 teeth in its jaws, spanning 5 rows. (See "external links" below)
Paleontologists suggest that a very large C. megalodon had jaws over 2 metres (7 ft) across.[11]
Bite force
In 2008, a team of scientists led by Dr. Stephen Wroe conducted an experiment to determine the bite force of C. megalodon; results indicate that it had one of the most powerful bites in history.[2] At 15.9 metres (52 ft) long, C. megalodon was capable of exerting a bite force estimated at 108,514 newtons (24,395 lbf),[2] and at 20.3 metres (67 ft) long, C. megalodon was capable of exerting a bite force estimated at 182,201 newtons (40,960 lbf).[2]
C. megalodon's bite force, at maximum estimated size, is over 28 times greater than that of Dunkleosteus at 5.3 kilonewtons (1,200 lbf),[2][29] over 10 times greater than that of the great white shark at 18 kilonewtons (4,000 lbf),[2] over 5 times greater than that of T. rex at 31 kilonewtons (7,000 lbf),[2] and also greater than that of Predator X at 150 kilonewtons (34,000 lbf).
In addition, Wroe and colleagues (2008) pointed out that sharks also shake sideways while feeding, amplifying the postcranial generated forces. Therefore the total forces experienced by prey are likely higher than the forces estimated through the experiment.[2] The extraordinary bite forces in C. megalodon must be considered in the context of the great size of this fossil predator and of paleontological evidence suggesting that C. megalodon was an active predator of large whales.[2]
Functional parameters of teeth
The exceptionally robust teeth of C. megalodon are serrated,[5][11] which would have improved efficiency in slicing the flesh of prey items. Paleontologist Dr. Bretton K. Kent from University of Maryland suggests that these teeth are comparatively thicker for their size with much lower slenderness and bending strength ratios. They also have roots that are substantially larger relative to total tooth heights, and so have a greater mechanical advantage. Teeth with these traits are not just good cutting tools but also are well suited for grasping powerful prey and would seldom crack even when slicing through bones.[30]
Skeletal anatomy
 Reconstructed Megalodon skeleton on display at the Calvert Marine Museum.
Reconstructed Megalodon skeleton on display at the Calvert Marine Museum.Aside from estimating the size of C. megalodon, Gottfried and colleagues (1996) also have tried to determine the schematics of the entire skeleton of C. megalodon.[7]
Jaw structure
To functionally support the very large and robust dentition, the jaws of the C. megalodon would have been massive, stouter, and more strongly developed than that of the great white shark, which possesses a somewhat gracile dentition in comparison.[7] The strongly developed jaws would have somewhat of a pig-eyed appearance.[7]
Chondrocranium
The chondrocranium of C. megalodon would have a blockier and more robust appearance than that of the great white shark, in order to functionally reflect its more massive jaws and dentition in comparison.[7]
Fins
The fins of C. megalodon would have been most likely proportionally larger and thicker in comparison to the fins of great white sharks because relatively larger fins are a necessity for propulsion and control of movements of a larger shark.[7]
Axial skeleton
Through thorough scrutiny of the partially preserved vertebral C. megalodon specimen from Belgium, it became apparent that C. megalodon had a higher vertebral count than found in large specimens of any known shark.[7] Only the vertebral count in great white shark came close in quantity, symbolizing close anatomical ties between the two species.[7]
Full skeletal reconstruction
On the basis of the characteristics mentioned above, Gottfried and colleagues (1996) eventually managed to reconstruct the entire skeleton of C. megalodon, which has been put on display in Calvert Marine Museum at Solomons island, Maryland in USA.[7][31][32] This C. megalodon skeletal reconstruction is 11.5 metres (38 ft) long and represents a young individual.[7] The team stresses that relative and proportional changes in the skeletal features of C. megalodon are ontogenetic in nature in comparison to that of great white shark, as they occur in great white sharks while growing larger.[7] In addition, the fossil remains of C. megalodon confirm that it had a heavily calcified skeleton in life.[12]
Paleoecological considerations
Range and habitat
Sharks, especially large species, are highly mobile organisms with a complex life history and wide distribution.[1] Fossil records of C. megalodon indicate that it was cosmopolitan,[11] and commonly occurred in subtropical to temperate latitudes.[7] Prior to the formation of the Isthmus of Panama, the seas were relatively warmer.[33] This would have made it possible for the species to live in all the oceans of the world.
C. megalodon had enough behavioral flexibility to inhabit wide range of marine environments (i.e. coastal shallow waters,[34] coastal upwelling,[34] swampy coastal lagoons,[34] sandy littorals,[34] and offshore deep water environments[11]), and exhibited a transient life-style.[34] Adult C. megalodon were not abundant in shallow water environments,[34] and mostly lurked offshore. C. megalodon may have moved between coastal and oceanic waters, particularly in different stages in its life cycle.
Prey relationships
 Vertebral centrum of a whale bitten in half by a megalodon. Large bite marks (deep gashes) on the vertebral centrum are visible.
Vertebral centrum of a whale bitten in half by a megalodon. Large bite marks (deep gashes) on the vertebral centrum are visible.Sharks are generally opportunistic predators. However, scientists propose that C. megalodon was "arguably the most formidable carnivore ever to have existed."[2] Its great size,[2] high-speed swimming capability,[32] and powerful jaws coupled with formidable killing apparatus,[2][7] made it a super-predator with the capability to consume a broad spectrum of fauna.
Fossil evidence indicates that C. megalodon preyed upon cetaceans (i.e., dolphins,[7] small whales,[4] (including cetotherrids,[10] squalodontids,[34] and Odobenocetops[35]), and large whales,[36] (including sperm whales,[11][37] bowhead whales,[38] and rorquals[36][39]), pinnipeds,[10][17][40] porpoises,[11] sirenians,[34][41] and giant sea turtles.[34]
Marine mammals were regular prey targets for C. megalodon. Many whale bones have been found with clear signs of large bite marks (deep gashes) made by teeth that match those of C. megalodon,[4][7] and various excavations have revealed C. megalodon teeth lying close to the chewed remains of whales,[7][31] and sometimes in direct association with them.[8] Fossil evidence of interactions between C. megalodon and pinnipeds also exist.[17] In one interesting observation, a 127 millimetres (5.0 in) C. megalodon tooth was found lying very close to a bitten earbone of a sea lion.[40]
 Artistic impression of a Megalodon pursuing two Eobalaenoptera whales.
Artistic impression of a Megalodon pursuing two Eobalaenoptera whales.C. megalodon faced a highly competitive environment during its time of existence.[42] However, C. megalodon, being at the top of the food chain,[43] likely had a profound impact on the structuring of marine communities.[7][42][44] Fossil evidence indicates a correlation between the emergence of C. megalodon and extensive diversification of cetaceans around the world.[7][42] Juvenile C. megalodon preferred regions where small cetaceans were abundant, and adult C. megalodon preferred regions where large cetaceans were abundant.[7] Such preferences may have developed shortly after they appeared in the Oligocene.[7] In addition, C. megalodon was contemporaneous with macro-predatory odontocetes (particularly raptorial sperm whales and squalodontids), which were also likely among the apex predators of that time,[9][42] and provided competition.[9][45] In response to competition from giant macro-predatory sharks, macro-predatory odontocetes may have evolved some defensive adaptations; some species became pack predators,[9][46] and some species attained gigantic sizes, such as Livyatan melvillei.[42][47] By the end of the Miocene, raptorial sperm whales vanished from the fossil record and left an ecological void.[42]
Like other sharks, C. megalodon also would have been piscivorous.[10][32] Fossil evidence indicates that other notable species of macro-predatory sharks (e.g. great white sharks) responded to competitive pressure from C. megalodon by avoiding regions it inhabited.[7] C. megalodon likely also had a tendency for cannibalism.[48]
Feeding strategies
Sharks often employ complex hunting strategies to engage large prey animals. Some paleontologists suggest that the hunting strategies of the great white shark may offer clues as to how C. megalodon might have hunted its unusually large prey (i.e., whales).[9] However, fossil evidence suggests that C. megalodon employed more effective hunting strategies against large prey than those of the great white shark.[30]
Paleontologists have conducted a survey of fossils to determine attacking patterns of C. megalodon on prey.[30] One particular specimen — the remains of a 9 metres (30 ft) long prehistoric baleen whale (of an unknown Miocene taxon) — provided the first opportunity to quantitatively analyze the attacking behavior of C. megalodon.[30] The predator primarily focused its attack on the tough bony portions (i.e. shoulders, flippers, rib cage, and upper spine) of the prey,[30] which great white sharks generally avoid.[30] Dr. Bretton Kent elaborated that C. megalodon attempted to crush the bones and damage delicate organs (i.e. heart, and lungs) harbored within the rib cage of the prey.[30] Such an attack would have immobilized the prey, which would have died quickly due to injuries to these vital organs.[30] These findings also clarify why the ancient shark needed more robust dentition than that of great white sharks.[30] Furthermore, the attack patterns could differ for prey of different sizes.[9] Fossil remains of some small cetaceans (e.g. cetotheriids) suggest that they were rammed with great force from below before being killed and eaten.[9]
During the Pliocene, larger and and more advanced cetaceans appeared.[49] C. megalodon apparently further refined its hunting strategies to cope with these large whales. Numerous fossilized flipper bones (i.e., segments of the pectoral fins), and caudal vertebrae of large whales from the Pliocene have been found with bite marks that were caused by attacks of C. megalodon.[2] This paleontological evidence suggests that C. megalodon would attempt to immobilize a large whale by ripping apart or biting off its propulsive structures before killing and feeding on it.[2]
Nursery areas
 Collection of teeth of juvenile Megalodon from a probable nursery area in the Gatun Formation of Panama.
Collection of teeth of juvenile Megalodon from a probable nursery area in the Gatun Formation of Panama.Fossil evidence suggests that the preferred nursery sites of C. megalodon were warm water coastal environments, where potential threats were minor and food sources plentiful.[1] Nursery sites have been identified in the Gatun Formation of Panama,[1] the Calvert Formation of Maryland,[1] and the Bone Valley Formation of Florida.[1] As is the case with most sharks, C. megalodon also gave birth to live young. The size of neonate C. megalodon teeth indicate that C. megalodon pups were around 2 to 4 metres (7 to 13 ft) in total length (TL) at birth.[1][7] Their dietary preferences display an ontogenetic shift.[7] Young C. megalodon commonly preyed on fish,[1] giant sea turtles,[34] dugongs,[11] and small cetaceans;[7] mature C. megalodon moved to off-shore cetacean high-use areas and consumed large cetaceans.[7]
However, there is an exceptional case which suggests that juvenile C. megalodon may occasionally have attacked much larger balaenopterid whales. Three tooth marks apparently from a 4-7 m long Pliocene macro-predatory shark were found on a rib from an ancestral great blue or humpback whale that showed evidence of subsequent healing.[39][50][51] Scientists suspect that this shark was a juvenile C. megalodon.[50][51]
Extinction
The subject of the extinction of C. megalodon is still under investigation.[9] Several possible causes for its decline and eventual disappearance have been proposed.
Oceanic cooling and sea level drops
The closure of Central American Seaway fundamentally changed global ocean circulation.[4][52] This event initially set the stage for glaciation in the northern hemisphere,[52] and later on, also facilitated the cooling of the entire planet.[52] In addition, expansion of glaciation during the Pliocene tied up huge volumes of water in the form of ice sheets, resulting in significant sea level drops.[33] Consequently, during the late Pliocene and Pleistocene, there were ice ages,[53][54] which cooled the oceans significantly.[4] The major reason cited behind the extinction of C. megalodon is the decline in ocean temperatures at global scale during the Pliocene.[4][31][55] This cooling trend adversely impacted C. megalodon, as it preferred warmer waters,[7][55] and as a result it became extremely rare until its ultimate extinction during the Pleistocene.[55] Fossil evidence confirms the absence of C. megalodon in regions around the world where water temperatures had significantly declined during the Pliocene.[7] Furthermore, these oceanographic changes may have restricted many of the suitable warm water nursery sites for C. megalodon, hindering population maintenance.[4] Nursery areas are pivotal for the survival of a species.[56]
Decline in food supply
Cetaceans attained their greatest diversity during the Miocene,[7] with over 20 recognized genera in comparison to only six living genera.[57] Such diversity presented an ideal setting to support a gigantic macro-predator like C. megalodon.[7] However, by the end-Miocene, many species of cetaceans became extinct.[42] Furthermore, after the closure of Central American Seaway, additional extinctions occurred in the marine environment,[58][59] and faunal redistribution took place; the great whales abandoned the tropics.[60] Whale migratory patterns during the Pliocene have been reconstructed from the fossil record, suggesting that most surviving species showed a trend towards polar regions.[33] The cooling trend in oceans during the Pliocene prevented C. megalodon from gaining access to polar regions, depriving the gigantic shark from its main food source; the great whales.[4][7][10][60] As a result of these developments, the food supply for C. megalodon in regions it inhabited during the Pliocene, primarily in low-to-mid latitudes, was no longer sufficient to sustain the species worldwide.[4][36][60] C. megalodon was adapted to a specialized lifestyle,[31] and this lifestyle was disturbed as a result of these developments.[31] In addition, the resulting shortage of food sources in tropics during Plio-Pleistocene times may have fueled cannibalism within C. megalodon.[9] The juvenile individuals were at increased risk from attacks by adult individuals during times of starvation.[9]
New competition
Large raptorial delphinids (members of genus Orcinus) evolved during the Pliocene,[61] and likely filled the ecological void left by the disappearance of raptorial sperm whales at the end of the Miocene.[42] A minority view is that these delphinids may have outcompeted the last representatives of C. megalodon for the top predator niche.[62] Fossil records indicate that these delphinids commonly occurred at high latitudes during the Pliocene,[61] indicating that they could cope with the increasingly prevalent cold water temperatures.[61] These delphinids also occurred in tropics (e.g. Orcinus sp. in South Africa).[61] Large bite marks on the fossil remains of delphinids indicate that C. megalodon preyed on them.[7] However, Paleontologist Albert Sanders suggests that C. megalodon had become too large to sustain itself on the available food supply in the tropics.[56] Most experts have inferred that factors such as cooling trend in oceans, and shortage of food sources, during Plio-Pleistocene times have played a significant role in downfall and demise of C. megalodon.[9] Other apex predators seem to have gained from the extinction of this formidable species.[55]
Taxonomy
Even after decades of research and scrutiny, controversy over the phylogeny of C. megalodon still persists.[5][63] Several shark researchers (e.g. J. E. Randall, A. P. Klimley, D. G. Ainley, M. D. Gottfried, L. J. V. Compagno, S. C. Bowman, and R. W. Purdy) insist that C. megalodon is a close relative of the great white shark. However, several other shark researchers (e.g. D. S. Jordan, H. Hannibal, E. Casier, C. DeMuizon, T. J. DeVries, D. Ward, and H. Cappetta) dismiss the proposal that C. megalodon is a close relative of the great white shark, and cite convergent evolution as the reason for the dental similarity. The arguments of the supporters of the Carcharocles genus for C. megalodon seem to have gained noticeable support.[31] However, the original taxonomic assignment still has wide acceptance.[5]
Megalodon within Carcharodon
The megalodon hypothesis claims that C. carcharias is more closely related to C. megalodon than I. hastalis.[5]Lamniformes I. hastalis
Sacaco sp.
C. carcharias
C. megalodon
The Otodus-Origin hypothesis claims that C. carcharias descends from megatoothed sharks.[63]Lamniformes O. obliquus
I. hastalis
C. carcharias
C. megalodon
The traditional view is that C. megalodon should be classified within the genus Carcharodon along with the great white shark. Main reasons cited for this phylogeny are; (1) an ontogenetic gradation, whereby the teeth of C. carcharias shift from having coarse serrations as a juvenile to fine serrations as an adult, the latter resemble those of C. megalodon; (2) morphological similarity of teeth of young C. megalodon to those of C. carcharias; (3) a symmetrical second anterior tooth; (4) large intermediate tooth that is inclined mesially; and (5) upper anterior teeth that have a chevron-shaped neck area on the lingual surface. The supporters of classification as Carcharodon for C. megalodon suggest that C. megalodon and C. carcharias share a common ancestor, Palaeocarcharodon orientalis.[5][11]
Megalodon within Carcharocles
The hastalis hypothesis claims that C. carcharias is more closely related to I. hastalis than to C. megalodon.[5]Lamniformes I. hastalis
Sacaco sp.
C. carcharias
void C. megalodon
The Isurus-Origin hypothesis claims that C. carcharias descends from I. hastalis.[63]Lamniformes O. obliquus
C. megalodon
I. hastalis
C. carcharias
Around 1923, the genus Carcharocles was proposed by two shark researchers, D. S. Jordan and H. Hannibal, to classify a shark C. auriculatus. Later on, Carcharocles proponents assigned C. megalodon to Carcharocles genus.[5][11] Carcharocles proponents also suggest that the direct ancestor of the sharks belonging to the Carcharocles genus, is an ancient giant shark called Otodus obliquus, which lived during the Paleocene and Eocene epochs.[31][63] According to supporters of classification as Carcharocles for C. megalodon; Otodus obliquus evolved in to Otodus aksuaticus,[11][31] which evolved in to Carcharocles auriculatus,[11][31] which evolved into Carcharocles angustidens,[11][31] which evolved into Carcharocles chubutensis,[11][31] which eventually evolved into megalodon.[11][31] Hence, the immediate ancestor of C. megalodon is Carcharocles chubutensis,[11][31] because it serves as the missing link between Carcharocles augustidens and C. megalodon and it bridges the loss of the "lateral cusps" that characterize C. megalodon.[11][31]
Megalodon as chronospecies?
Shark researcher David Ward has further elaborated on the Carcharocles evolutionary process by implying that this lineage, stretching from the Paleocene to the Pliocene, is of a single giant shark which gradually changed through time, suggesting a case of chronospecies.[11]
New evolutionary position for great white shark
Carcharocles proponents point out that the great white shark is closely related to an ancient shark Isurus hastalis, the "broad tooth mako", rather than to C. megalodon.[5] One reason cited by paleontologist Dr. Chuck Ciampaglio is that the dental morphometrics (variations and changes in the physical form of objects) of I. hastalis and C. carcharias are remarkably similar.[5] Another reason cited is that C. megalodon teeth have much finer serrations than in C. carcharias teeth.[5] Further evidence linking the great white shark more closely to ancient mako sharks, rather than to C. megalodon, has been provided in 2009 — The fossilized remains of an ancient form of the great white shark were excavated from southwestern Peru in 1988, which are about 4 million years old. These fossilized remains demonstrate a likely shared ancestor of modern mako and great white sharks.[63][64]
Existing controversy and considerations
 Two teeth of great white shark (white) compared to a single tooth of Megalodon (black). A coin 23 mm in width shows the scale.
Two teeth of great white shark (white) compared to a single tooth of Megalodon (black). A coin 23 mm in width shows the scale.Paleontologist Dr. Chuck Ciampaglio from Wright State University asserts that similarities between the teeth of C. megalodon and the great white shark are superficial and there are noticeable morphometric differences between them, and that these findings are sufficient to warrant a separate genus for C. megalodon.[5][9] However, some proponents of the Carcharodon genus for C. megalodon (i.e. M. D. Gottfried, and R. E. Fordyce) have provided more arguments for a close relationship between the extinct megatooth sharks and the great white shark.[19] With respect to the recent controversy regarding fossil lamnid shark relationships, the overall morphology – particularly the internal calcification patterns – of the great white shark vertebral centra have been compared to well-preserved fossil centra from the megatooth sharks, including C. megalodon and C. angustidens. The morphological similarity apparent from these comparisons supports a close relationship of the giant fossil megatooth species to living white sharks.[19][65]
With respect to the origins of the great white shark, Gottfried and Fordyce have pointed out that some great white shark fossils are about 16 million years old and predate the transitional Pliocene fossils.[19] In addition, the Oligocene records of C. megalodon,[11][19] contradict the suggestion that Carcharocles chubutensis is the immediate ancestor of C. megalodon. These records also indicate that C. megalodon actually co-existed with Carcharocles angustidens.[19] Hence, proponents for the Carcharodon genus for C. megalodon argue that extinct megatoothed sharks should be placed within the genus Carcharodon.[19]
Some paleontologists argue that the genus Otodus should be used for sharks within the Carcharocles lineage and the genus Carcharocles should be discarded.[1]
At present, several proponents of the Carcharocles genus for C. megalodon (i.e. Catalina Pimiento, Dana J. Ehret, Bruce J. MacFadden, and Gordon Hubbell) accept that both species belong to the order Lamniformes, and in the absence of living members of the family Otodontidae, great white shark is the most ecologically analogous species to C. megalodon.[1]
In fiction
Ever since the remains of C. megalodon were discovered, it has been an object of fascination. It has been portrayed in several works of fiction, including films and novels, and continues to hold its place among the most popular subjects for fiction involving sea monsters. Many of these works of fiction posit that at least a relict population of C. megalodon survived extinction and lurk in the vast depths of the ocean, and that individuals may manage to surface from the vast depths, either by human intervention or by natural means. Jim Shepard's story "Tedford and the Megalodon" is a good example of this. Such beliefs are usually inspired by the discovery of a C. megalodon tooth by members of the HMS Challenger in 1872, which some believed to be only 10,000 years old.[66] However, this tooth has been re-examined, and findings indicate that it is untestable for age.[66]
Some works of fiction (such as Shark Attack 3: Megalodon and Steve Alten's Meg series) incorrectly depict C. megalodon as being a species over 70 million years old, and to have been alive during the time of dinosaurs. The writers of the movie Shark Attack 3: Megalodon depicted this assumption by including an altered copy of a book by the shark researcher, Richard Ellis, called "Great White Shark". The copy shown in the film had several pages that do not exist in the real book. The author of the real book sued the film's distributor Lions Gate Entertainment, asking for a halt to the film's distribution along with $150,000 in damages.[67] Steve Alten's Meg: A Novel of Deep Terror is probably best known for portraying this inaccuracy with its prologue and cover artworks depicting C. megalodon killing a tyrannosaur in the sea.
Mega Shark Versus Giant Octopus (2009) and its sequel Mega Shark Versus Crocosaurus (2010) are parodies of disaster films with a giant shark similar to a megalodon in the title role.
See also
- For a topical guide to this subject, see Outline of sharks.
- List of prehistoric cartilaginous fish
- Prehistoric fish
- Largest prehistoric organisms
Footnotes
- ^ a b c d e f g h i j k l m n o p q r s t u v w Pimiento, Catalina; Dana J. Ehret, Bruce J. MacFadden, and Gordon Hubbell (May 10, 2010). Stepanova, Anna. ed. "Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama". PLoS One (Panama: PLoS.org) 5 (5): e10552. doi:10.1371/journal.pone.0010552. PMC 2866656. PMID 20479893. http://www.plosone.org/article/info%3Adoi%2F10.1371%2Fjournal.pone.0010552. Retrieved 12 May 2010.
- ^ a b c d e f g h i j k l m n o p q r Wroe, S.; Huber, D. R. ; Lowry, M. ; McHenry, C. ; Moreno, K. ; Clausen, P. ; Ferrara, T. L. ; Cunningham, E. ; Dean, M. N. ; Summers, A. P. (2008). "Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?". Journal of Zoology 276 (4): 336–342. doi:10.1111/j.1469-7998.2008.00494.x. http://www.bio-nica.info/Biblioteca/Wroe2008GreatWhiteSharkBiteForce.pdf.
- ^ Haven, Kendall (1997). 100 Greatest Science Discoveries of All Time. Libraries Unlimited. pp. 25–26. ISBN 1591582652.
- ^ a b c d e f g h i j k l m n o p q r s Bruner, J. C. (Sept.-Oct. 1997). "The Megatooth shark, Carcharodon megalodon: Rough toothed, huge toothed". Mundo Marino Revista Internacional de Vida (non-refereed) (Marina) 5: 6-11. http://www.sharksteeth.com/megatoothshark.htm. Retrieved 2011-11-14.
- ^ a b c d e f g h i j k l m n Nyberg K.G, Ciampaglio C.N, Wray G.A (2006). "Tracing the ancestry of the GREAT WHITE SHARK". Journal of Vertebrate Paleontology 26 (4): 806–814. doi:10.1671/0272-4634(2006)26[806:TTAOTG]2.0.CO;2. http://www.bioone.org/doi/abs/10.1671/0272-4634(2006)26%5B806:TTAOTG%5D2.0.CO%3B2. Retrieved 2007-12-25.
- ^ Agassiz, Louis (1833-1843). Recherches sur les poissons fossiles ... / par Louis Agassiz.. Neuchatel :Petitpierre. p. 41. http://www.biodiversitylibrary.org/bibliography/4275. Retrieved 2008-09-08.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf Klimley, Peter; Ainley, David (1996). Great White Sharks: The Biology of Carcharodon carcharias. Academic Press. ISBN 0124150314. http://www.elsevier.com/wps/find/bookdescription.cws_home/673659/description#description.
- ^ a b Augilera, Orangel A.; Garcia, Luis; Cozzoul, Mario A. (2008). "Giant-toothed white sharks and cetacean trophic interaction from the Pliocene Caribbean Paraguaná Formation". Paläontologische Zeitschrift (Springer Berlin) 82 (2): 204–208. doi:10.1007/BF02988410 (inactive 2010-03-17). http://www.springerlink.com/content/637327q841643568/.
- ^ a b c d e f g h i j k l "Monster Shark". Narrated by: Robert Leigh. Prehistoric Predators. National Geographic. 2009-04-27.
- ^ a b c d e f Roesch, Ben (1998). "The Cryptozoology Review: A Critical Evaluation of the Supposed Contemporary Existence of Carcharocles Megalodon". http://www.ncf.carleton.ca/~bz050/megalodon.html.
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad Renz, Mark (2002). Megalodon: Hunting the Hunter. PaleoPress. ISBN 0-9719477-0-8. http://google.com/books?id=cMRe5GmDXmUC&printsec=frontcover.
- ^ a b Bendix-Almgreen, Svend Erik (November 15, 1983). "Carcharodon megalodon from the Upper Miocene of Denmark, with comments on elasmobranch tooth enameloid: coronoi'n". Bulletin of the Geological Society of Denmark (Copenhagen: Geologisk Museum) 32: 1–32. http://2dgf.dk/xpdf/bull32-01-02-1-32.pdf. Retrieved March 2010.
- ^ Nieves-Rivera, Angel M.; Ruizyantin, Maria ; Gottfried, Michael D. (2003). "New Record of the Lamnid Shark Carcharodon megalodon from the Middle Miocene of Puerto Rico". Caribbean Journal of Science 39: 223–227.
- ^ Iturralde-Vinent, M.; G. Hubbell, and R. Rojas (1996). "CATALOG OF CUBAN FOSSIL ELASMOBRANCHII (PALEOCENE--PLIOCENE) AND PALEOOCEANOGRAPHIC IMPLICATIONS OF THEIR LOWER--MIDDLE MIOCENE OCCURRENCE". Boletín de la Sociedad Jamaicana de Geología (Cuba) 31: 7–21. http://www.redciencia.cu/cdorigen/arca/paper/cubff.pdf. Retrieved 10 May 2010.
- ^ Donovan, Stephen; Gavin, Gunter (2001). Fossil sharks from Jamaica. 28. Bulletin of the Mizunami Fossil Museum. pp. 211–215. http://www.city.mizunami.gifu.jp/odocs/sightseeing/mizunami/cultural_property/institution/Bull28/BMFM28-211-215(148kB).pdf.
- ^ Fitzgerald, Erich (2004). "A review of the Tertiary fossil Cetacea (Mammalia) localities in Australia". Memoirs of Museum Victoria (Australia: Museum Victoria) 61 (2): 183–208. ISSN 1447-2554. http://museumvictoria.com.au/pages/3948/61_2_fitzgerald.pdf. Retrieved March 2010.
- ^ a b c Portell, Roger; Hubell, Gordon; Donovan, Stephen; Green, Jeremy; Harper, David; Pickerill, Ron (2008). Miocene sharks in the Kendeace and Grand Bay formations of Carriacou, The Grenadines, Lesser Antilles. 44. Caribbean Journal of Science. pp. 279–286. http://caribjsci.org/Dec08/44_279-286.pdf.
- ^ Hideo, Habe; Mastatoshi, Goto; Naotomo, Kaneko (22 March 2004). "Age of Carcharocles megalodon (Lamniformes: Otodontidae) : A review of the stratigraphic records". The Palaeontological Society of Japan (PSJ) (Japan) 75: 7–15. ISSN 0022-9202. http://ci.nii.ac.jp/naid/110007574397.
- ^ a b c d e f g h i Gottfried M.D., Fordyce R.E. (2001). "An Associated Specimen of CARCHARODON ANGUSTIDENS (CHONDRICHTHYES, LAMNIDAE) From the LATE OLIGOCENE of NEW ZEALAND, with comments on CARCHARODON Interrelationships". Journal of Vertebrate Paleontology 21 (4): 730–739. doi:10.1671/0272-4634(2001)021[0730:AASOCA]2.0.CO;2. ISSN 0272-4634.
- ^ Brown, Robin (2008). Florida's Fossils. Pineapple Press. ISBN 978-1-56164-409-4.
- ^ a b c d e f g Helfman, Gene; Collette, Bruce; Facey, Douglas (1997). The diversity of fishes. Wiley Blackwell. ISBN 978-0-8654-2256-8.
- ^ a b c d e Randall, John (July 1973). "Size of the Great White Shark (Carcharodon)". Science Magazine: 169–170.
- ^ Kowinsky, Jayson (2002). "The Size of Megalodons". http://www.fossilguy.com/topics/megsize/megsize.htm. Retrieved 2008-01-12.
- ^ a b c d e f g [|Shimada, Kenshu] (5 Nov 2002). "The relationship between the tooth size and total body length in the white shark, Carcharodon carcharias (Lamniformes: Lamnidae)". Journal of Fossil Research (Japan) 35 (2): 28–33. ISSN 0387-1924. http://sciencelinks.jp/j-east/article/200309/000020030903A0178835.php. Retrieved April 29, 2011.[dead link]
- ^ Schembri, Patrick (1994). "MALTA'S NATURAL HERITAGE". Natural Heritage. in (MALTA: University of MALTA): 105–124. http://home.um.edu.mt/biology/PJS%20researches/Malta's_natural_heritage_(1994).pdf. Retrieved March 2010.
- ^ Papson, Stephen (1992). "Copyright: Cross the Fin Line of Terror". Journal of American Culture 15: 67–81
- ^ a b Compagno, Leonard J. V. (2002). SHARKS OF THE WORLD: An Annotated and Illustrated Catalogue of Shark Species Known to Date. Rome: Food & Agriculture Organization of the United Nations. p. 97. ISBN 9251045437.
- ^ a b c Herskowitz, David; Peter Wiggins (21 December 2010). "Huge Shark Jaw" (PDF). Natural History. Heritage Auctions. http://fineart.ha.com/s/d/shark-jaws.pdf. Retrieved April 30, 2011.
- ^ Anderson, Philip; Westneat, Mark (22). "Feeding mechanics and bite force modelling of the skull of Dunkleosteus terrelli, an ancient apex predator". Royal Society 3 (1): 77–80. doi:10.1098/rsbl.2006.0569. http://rsbl.royalsocietypublishing.org/content/3/1/77.full
- ^ a b c d e f g h i Riordon, James (June 1999). "Hell's teeth". NewScientist Magazine (2190): 32. http://www.newscientist.com/article/mg16221904.900-hells-teeth.html.
- ^ a b c d e f g h i j k l m n Andres, Lutz (2002). "C. megalodon — Megatooth Shark, Carcharodon versus Carcharocles". http://www.fossilguy.com/topics/megshark/megshark.htm. Retrieved 2008-01-16.
- ^ a b c Arnold, Caroline (2000). Giant Shark: Megalodon, Prehistoric Super Predator. Houghton Mifflin. pp. 18–19. ISBN 9780395914199.
- ^ a b c Gillette, Lynett. "Winds of Change". San Diego Natural History Museum. http://www.sdnhm.org/research/readings/fn_0307.html. Retrieved 2009-09-25.
- ^ a b c d e f g h i j Aguilera O., Augilera E. R. D. (2004). "Giant-toothed White Sharks and Wide-toothed Mako (Lamnidae) from the Venezuela Neogene: Their Role in the Caribbean, Shallow-water Fish Assemblage". Caribbean Journal of Science 40 (3): 362–368.
- ^ "Fact File: Odobenocetops". BBC. http://www.bbc.co.uk/science/seamonsters/factfiles/odobenocetops.shtml. Retrieved 2008-03-21.
- ^ a b c Morgan, Gary S. (August 1994). "Whither the giant white shark?". Paleontology Topics. Paleontological Research Institution.
- ^ "MEGALODON". Fossil Farm Museum Of The Fingerlakes. http://fingerlakesfossilfarm.org/mammal_fossils.htm. Retrieved 2010-07-01.
- ^ deGruy, Michael (2006). Perfect Shark (TV-Series). BBC.
- ^ a b Godfrey, Stephen (April 2004). "The Ecphora: Fascinating Fossil Finds" (PDF). Paleontology Topics. Calvert Marine Museum. http://www.calvertmarinemuseum.com/cmmfc/newsletter/CMMFC_Newsletter_2004-04.pdf. Retrieved 21 February 2010.
- ^ a b Kehe, Andy. "Bone Apetite". http://www.sharktoothhill.org/bone.html. Retrieved 17 February 2011.
- ^ Godfrey, Stephen (March 2007). "The Ecphora: Shark-Bitten Sea Cow Rib" (PDF). Paleontology Topics. Calvert Marine Museum. http://www.calvertmarinemuseum.com/cmmfc/newsletter/CMMFC_Newsletter_2007-03.pdf. Retrieved 27 March 2010.
- ^ a b c d e f g h Lambert, Olivier; Giovanni Bianucci, Klaas Post, Christian de Muizon, Rodolfo Salas-Gismondi, Mario Urbina and Jelle Reumer (1 July 2010). "The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru". Nature (Peru) 466 (7302): 105–108. doi:10.1038/nature09067. PMID 20596020. http://www.nature.com/nature/journal/v466/n7302/full/nature09067.html.
- ^ Compagno, Leonard J. V. (May 1989). "Copyright: Alternative life-history styles of cartilaginous fishes in time and space". Environmental Biology of Fishes 28: 33–75. doi:10.1007/BF00751027
- ^ Ferretti, Francesco; Boris Worm, Gregory L. Britten, Michael R. Heithaus, and Heike K. Lotze1 (August 2010). "Patterns and ecosystem consequences of shark declines in the ocean". Ecology Letters (Blackwell Publishing Ltd) 13 (8): 1055–1071. doi:10.1111/j.1461-0248.2010.01489.x. PMID 20528897. http://wormlab.biology.dal.ca/ramweb/papers-total/Ferretti%20et%20al%202010_Ecol_Lett.pdf. Retrieved 19 February 2011.
- ^ "New Leviathan Whale Was Prehistoric "Jaws"?". National Geographic Daily News (Peru: National Geographic). 30 June 2010. http://news.nationalgeographic.com/news/2010/06/photogalleries/100630-leviathan-mellvillei-sperm-whale-fossils-science/#whale02-scientists-skull-desert_22738_600x450.jpg.
- ^ Bianucci, Giovanni; Walter, Landini (8 Sep 2006). "Killer sperm whale: a new basal physeteroid (Mammalia, Cetacea) from the Late Miocene of Italy". Zoological Journal of the Linnean Society 148 (1): 103–131. doi:10.1111/j.1096-3642.2006.00228.x
- ^ "Ancient monster whale more fearsome than Moby Dick". NewScientist. http://www.newscientist.com/article/dn19108-ancient-monster-whale-more-fearsome-than-moby-dick.html. Retrieved 2010-06-30.
- ^ Tanke, Darren; Currie, Philip (December 1998). Head-Biting Behaviour in Theropod Dinosaurs: Paleopathological Evidence. Gaia 15. p. 168. ISSN 0871-5424. http://www.mnhn.ul.pt/geologia/gaia/12.pdf.[dead link]
- ^ Deméré, Thomas A.; Berta, Annalisa; McGowen, Michael R. (2005). "The taxonomic and evolutionary history of fossil and modern balaenopteroid mysticetes". Journal of Mammalian Evolution 12 (1/2): 99–143. doi:10.1007/s10914-005-6944-3.
- ^ a b "Details of ancient shark attack preserved in fossil whale bone". Smithsonian. http://www.eurekalert.org/pub_releases/2011-11/s-doa111011.php. Retrieved 2011-11-12.
- ^ a b Kallal, R. J.; Godfrey, S. J.; Ortner, D. J. (2010-08-27). "Bone Reactions on a Pliocene Cetacean Rib Indicate Short-Term Survival of Predation Event". International Journal of Osteoarchaeology. doi:10.1002/oa.1199. http://onlinelibrary.wiley.com/doi/10.1002/oa.1199/abstract. Retrieved 2011-11-13.
- ^ a b c "How the Isthmus of Panama Put Ice in the Arctic". 2004-03-22. http://www.whoi.edu/oceanus/viewArticle.do?id=2508. Retrieved 2008-12-20.
- ^ "Pliocene epoch". http://www.ucmp.berkeley.edu/tertiary/pli.html. Retrieved 2008-01-16.
- ^ "Pleistocene epoch". http://www.ucmp.berkeley.edu/quaternary/ple.html. Retrieved 2008-01-16.
- ^ a b c d Antunes, Miguel Telles; Ausenda Cáceres Balbino (March 2010). "The Great White Shark Carcharodon carcharias (Linne, 1758) in the Pliocene of Portugal and its Early Distribution in Eastern Atlantic". Revista Española de Paleontología (Portugal) 25 (1): 1–6. ISSN 0213-6937. http://recyt.fecyt.es/index.php/REP/article/viewFile/11488/7741. Retrieved 19 February 2011.
- ^ a b Reilly, Michael (29 September 2009). "Prehistoric Shark Nursery Spawned Giants". USA: Discovery News. http://news.discovery.com/animals/megalodon-nursery-prehistoric-sharks.html.
- ^ Dooly A.C, Nicholas C.F, Luo Z.X (2004). "The Earliest known member of the RORQUAL". Journal of Vertebrate Paleontology 24 (2): 453–463. doi:10.1671/2401. http://www.seaturtle.org/PDF/Dooley_2004_JVertPaleo.pdf. Retrieved 2010-03-25.
- ^ Domning, Daryl P. (1 February 2001). "Sirenians, seagrasses, and Cenozoic ecological change in the Caribbean". Palaeogeography, Palaeoclimatology, Palaeoecology (USA: Elsevier) 166 (1–2): 27–50. doi:10.1016/S0031-0182(00)00200-5. http://foodweb.uhh.hawaii.edu/MARE390_files/Domning%202001.pdf. Retrieved 26 April 2011.
- ^ Fordyce, R. Ewan; Patrick G. Quilty, and James Daniels (2002). "Australodelphis mirus, a bizarre new toothless ziphiid-like fossil dolphin (Cetacea: Delphinidae) from the Pliocene of Vestfold Hills, East Antarctica". Antarctic Science (Cambridge University Press) 14 (1): 37–54. doi:10.1017/S0954102002000561.
- ^ a b c Allmon, Warren D.; Steven D. Emslie, Douglas S. Jones, and Gary S. Morgan (2006). "Late Neogene Oceanographic Change along Florida's West Coast: Evidence and Mechanisms". The Journal of Geology (USA: The University of Chicago) 104 (2): 143–162. Bibcode 1996JG....104..143A. doi:10.1086/629811.
- ^ a b c d Heyning, John; Dahlheim, Marilyn (15 January 1988). "Mammalian Species: Orcinus Orca". The American Society of Mammalogists 304: 1–9. http://www.science.smith.edu/departments/Biology/VHAYSSEN/msi/pdf/i0076-3519-304-01-0001.pdf
- ^ Turner, Pamela S. (Oct/Nov 2004). "Showdown at Sea: What happens when great white sharks go fin-to-fin with killer whales?". National Wildlife (National Wildlife Federation) 42 (6). http://www.nwf.org/nationalwildlife/article.cfm?issueid=70&articleid=991. Retrieved 2009-11-22.
- ^ a b c d e Ehret D. J., Hubbell G., Macfadden B. J. (2009). "Exceptional preservation of the white shark CARCHARODON from the early Pliocene of PERU". Journal of Vertebrate Paleontology 29 (1): 1–13. doi:10.1671/039.029.0113.
- ^ Dell-Amore, Christine (2009). "Most Complete Great White Fossil Yet". http://news.nationalgeographic.com/news/2009/03/090312-shark-jaw-picture.html. Retrieved 2009-03-16.
- ^ Godfrey, Stephen (November 11, 2006). "The Geology and Paleontology of Calvert Cliffs" (PDF). Paleontology Topics. The Ecphora Miscellaneous Publications. http://www.calvertmarinemuseum.com/cmmfc/Paleo-Symposium.pdf. Retrieved 2 November 2009.
- ^ a b "Mega Jaws". MonsterQuest. March 18, 2009. No. 7, season 3.
- ^ Boniello, Kathianne (2009-07-12). "Shark Film has Writer Biting Mad". New York Post. http://www.nypost.com/p/news/regional/shark_film_has_writer_biting_mad_1j0mXEU0EbAahqVeBpFaDL.
References
- Bretton W. Kent (1994). Fossil Sharks of the Chesapeake Bay Region. Egan Rees & Boyer, Inc. 146 pages. ISBN 1881620018
- Dickson K. A., Graham J. B. (2004). "Evolution and consequences of endothermy in fishes". Physiological and Biochemical Zoology 77 (6): 998–1018. doi:10.1086/423743. PMID 15674772.
External links
- Ancient Shark's Bite More Powerful Than T. Rex's from LiveScience
- Carcharocles: Extinct Megatoothed shark
- Fossil Field Guide, Carcharocles megalodon from San Diego Natural History Museum
- Fact File: Megalodon from BBC, with pictures and video
- Prehistoric Megalodon Information
- Quick Facts about Megalodon: Largest Shark That Ever Lived!
- The largest modern Megalodon jaw reconstruction in the world (update) from Museum of Nature and Science
- Shark Tales featuring Megalodon with demonstration of the chewing action of the giant shark by Dr. Chuck Ciampaglio
- Jurassic Shark
- 3-D image of Great White Shark compared to Megalodon
- Prehistoric Jaws killed whales with single bite by Steve Farrar, science correspondent
- Ocean Portal: Megalodon comparative size illustration by The Smithsonian Institution
- Fossil remains of a large Sperm whale with C. megalodon bite marks (an example of predator-prey relationship between C. megalodon and Sperm whales)
Paleontological videos
NOTE: Flash Player is required to view the content below.
- A video clip of the Perfect Shark (2006) show from BBC (Presents fossil evidence of predator-prey relationships of Megalodon)
- Video Gallery containing video clips featuring Megalodon from Discovery Channel
- Paleontologist Mark Renz shows a huge Megalodon tooth (one of the largest ever discovered) on YouTube
- A video clip depicting aggressive interspecific interactions between Megalodon and a pod of killer odontoceti (B. shigensis) from History Channel
- Animated size comparison of Megalodon with great white shark, human, and school bus from North Carolina Museum of Natural Sciences on YouTube.
- Prehistoric Washington DC: Mega Shark from Discovery Channel (Depicts attacking strategies of Megalodon on prey)
- Shark Week Special on Megalodon with Pat McCarthy and John Babiarz on YouTube with comments on its extinction.
- Megalodon fossil teeth show evidence of 10-million-year-old shark nursery on YouTube
- Expert view: information about the Megalodon on YouTube (featuring Dana Ehret)
Categories:- Lamnidae
- Oligocene fish
- Miocene fish
- Pliocene fish
- Pleistocene fish
- Prehistoric sharks












{kind=link}
{kind=link}
{kind=link}
Wikimedia Foundation. 2010.