- Color vision
-
 White light shone onto a green surface is perceived as green by the human eye, and processed as such in the brain's visual cortex.
White light shone onto a green surface is perceived as green by the human eye, and processed as such in the brain's visual cortex.

Color vision is the capacity of an organism or machine to distinguish objects based on the wavelengths (or frequencies) of the light they reflect, emit, or transmit. Colors can be measured and quantified in various ways; indeed, a human's perception of colors is a subjective process whereby the brain responds to the stimuli that are produced when incoming light reacts with the several types of cone photoreceptors in the eye.
Contents
Wavelength and hue detection
Isaac Newton discovered that white light splits into its component colors when passed through a prism, but that if those bands of colored light pass through another and rejoin, they make a white beam. The characteristic colors are, from low to high frequency: red, orange, yellow, green, cyan, blue, violet. Sufficient differences in frequency give rise to a difference in perceived hue; the just noticeable difference in wavelength varies from about 1 nm in the blue-green and yellow wavelengths, to 10 nm and more in the red and blue. Though the eye can distinguish up to a few hundred hues, when those pure spectral colors are mixed together or diluted with white light, the number of distinguishable chromaticities can be quite high.
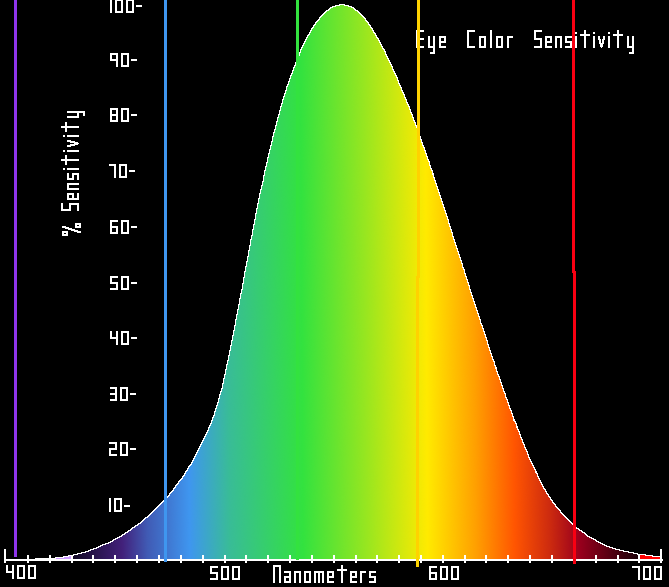
In very low light levels, vision is scotopic: light is detected by rod cells of the retina. Rods are maximally sensitive to wavelengths near 500 nm, and play little, if any, role in color vision. In brighter light, such as daylight, vision is photopic: light is detected by cone cells which are responsible for color vision. Cones are sensitive to a range of wavelengths, but are most sensitive to wavelengths near 555 nm. Between these regions, mesopic vision comes into play and both rods and cones provide signals to the retinal ganglion cells. The shift in color perception from dim light to daylight gives rise to differences known as the Purkinje effect.
The perception of "white" is formed by the entire spectrum of visible light, or by mixing colors of just a few wavelengths, such as red, green, and blue, or by mixing just a pair of complementary colors such as blue and yellow.[1]
Physiology of color perception
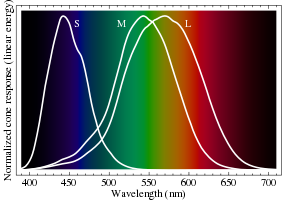 Normalized response spectra of human cones, S, M, and L types, to monochromatic spectral stimuli, with wavelength given in nanometers.
Normalized response spectra of human cones, S, M, and L types, to monochromatic spectral stimuli, with wavelength given in nanometers.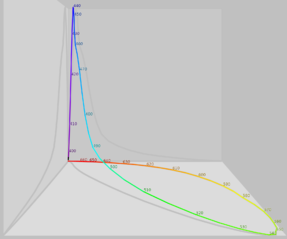 The same figures as above represented here as a single curve in three (normalized cone response) dimensions
The same figures as above represented here as a single curve in three (normalized cone response) dimensions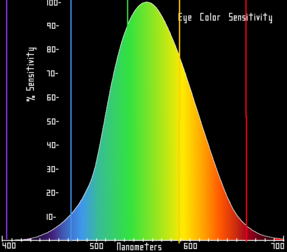 Relative brightness sensitivity of the human visual system as a function of wavelength
Relative brightness sensitivity of the human visual system as a function of wavelengthPerception of color begins with specialized retinal cells containing pigments with different spectral sensitivities, known as cone cells. In humans, there are three types of cones sensitive to three difference spectra, resulting in trichromatic color vision.
The cones are conventionally labeled according to the ordering of the wavelengths of the peaks of their spectral sensitivities: short (S), medium (M), and long (L) cone types. These three types do not correspond well to particular colors as we know them. Rather, the perception of color is achieved by a complex process that starts with the differential output of these cells in the retina and it will be finalized in the visual cortex and associative areas of the brain.
For example, while the L cones have been referred to simply as red receptors, microspectrophotometry has shown that their peak sensitivity is in the greenish-yellow region of the spectrum. Similarly, the S- and M-cones do not directly correspond to blue and green, although they are often depicted as such. It is important to note that the RGB color model is merely a convenient means for representing color, and is not directly based on the types of cones in the human eye.
The peak response of human cone cells varies, even among individuals with 'normal' color vision;[2] in non-human species this polymorphic variation is even greater, and it may well be adaptive.[3]
Theories of color vision
Two complementary theories of color vision are the trichromatic theory and the opponent process theory. The trichromatic theory, or Young–Helmholtz theory, proposed in the 19th century by Thomas Young and Hermann von Helmholtz, as mentioned above, states that the retina's three types of cones are preferentially sensitive to blue, green, and red. Ewald Hering proposed the opponent process theory in 1872.[4] It states that the visual system interprets color in an antagonistic way: red vs. green, blue vs. yellow, black vs. white. We now know both theories to be correct, describing different stages in visual physiology.[5]
Cone cells in the human eye
Cone type Name Range Peak wavelength[6][7] S β 400–500 nm 420–440 nm M γ 450–630 nm 534–555 nm L ρ 500–700 nm 564–580 nm A range of wavelengths of light stimulates each of these receptor types to varying degrees. Yellowish-green light, for example, stimulates both L and M cones equally strongly, but only stimulates S-cones weakly. Red light, on the other hand, stimulates L cones much more than M cones, and S cones hardly at all; blue-green light stimulates M cones more than L cones, and S cones a bit more strongly, and is also the peak stimulant for rod cells; and blue light stimulates S cones more strongly than red or green light, but L and M cones more weakly. The brain combines the information from each type of receptor to give rise to different perceptions of different wavelengths of light.
The opsins (photopigments) present in the L and M cones are encoded on the X chromosome; defective encoding of these leads to the two most common forms of color blindness. The OPN1LW gene, which codes for the opsin present in the L cones, is highly polymorphic (a recent study by Verrelli and Tishkoff found 85 variants in a sample of 236 men[8]). A very small percentage of women may have an extra type of color receptor because they have different alleles for the gene for the L opsin on each X chromosome. X chromosome inactivation means that only one opsin is expressed in each cone cell, and some women may therefore show a degree of tetrachromatic color vision.[9] Variations in OPN1MW, which codes the opsin expressed in M cones, appear to be rare, and the observed variants have no effect on spectral sensitivity.
Color in the human brain
 Visual pathways in the human brain. The ventral stream (purple) is important in color recognition. The dorsal stream (green) is also shown. They originate from a common source in the visual cortex.
Visual pathways in the human brain. The ventral stream (purple) is important in color recognition. The dorsal stream (green) is also shown. They originate from a common source in the visual cortex.Color processing begins at a very early level in the visual system (even within the retina) through initial color opponent mechanisms. Both Helmholtz's trichromatic theory, and Hering's opponent process theory are therefore correct, but trichromacy arises at the level of the receptors, and opponent processes arise at the level of retinal ganglion cells and beyond. In Hering's theory opponent mechanisms refer to the opposing color effect of red–green, blue–yellow, and light–dark. However, in the visual system, it is the activity of the different receptor types that are opposed. Some midget retinal ganglion cells oppose L and M cone activity, which corresponds loosely to red–green opponency, but actually runs along an axis from blue-green to magenta. Small bistratified retinal ganglion cells oppose input from the S cones to input from the L and M cones. This is often thought to correspond to blue–yellow opponency, but actually runs along a color axis from lime green to violet.
Visual information is then sent to the brain from retinal ganglion cells via the optic nerve to the optic chiasma: a point where the two optic nerves meet and information from the temporal (contralateral) visual field crosses to the other side of the brain. After the optic chiasma the visual tracts are referred to as the optic tracts, which enter the thalamus to synapse at the lateral geniculate nucleus (LGN).
The LGN is divided into laminae (zones), of which there three types: the M-laminae, consisting primarily of M-cells, the P-laminae, consisting primarily of P-cells, and the koniocellular laminae. M- and P-cells received relatively balanced input from both L- and M-cones throughout most of the retina, although this seems to not be the case at the fovea, with midget cells synapsing in the P-laminae. The koniocellular laminae receive axons from the small bistratified ganglion cells.[10][11]
After synapsing at the LGN, the visual tract continues on back to the primary visual cortex (V1) located at the back of the brain within the occipital lobe. Within V1 there is a distinct band (striation). This is also referred to as "striate cortex", with other cortical visual regions referred to collectively as "extrastriate cortex". It is at this stage that color processing becomes much more complicated.
In V1 the simple three-color segregation begins to break down. Many cells in V1 respond to some parts of the spectrum better than others, but this "color tuning" is often different depending on the adaptation state of the visual system. A given cell that might respond best to long wavelength light if the light is relatively bright might then become responsive to all wavelengths if the stimulus is relatively dim. Because the color tuning of these cells is not stable, some believe that a different, relatively small, population of neurons in V1 is responsible for color vision. These specialized "color cells" often have receptive fields that can compute local cone ratios. Such "double-opponent" cells were initially described in the goldfish retina by Nigel Daw;[12][13] their existence in primates was suggested by David H. Hubel and Torsten Wiesel and subsequently proven by Bevil Conway.[14] As Margaret Livingstone and David Hubel showed, double opponent cells are clustered within localized regions of V1 called blobs, and are thought to come in two flavors, red–green and blue–yellow.[15] Red–green cells compare the relative amounts of red–green in one part of a scene with the amount of red–green in an adjacent part of the scene, responding best to local color contrast (red next to green). Modeling studies have shown that double-opponent cells are ideal candidates for the neural machinery of color constancy explained by Edwin H. Land in his retinex theory.[16]
 This image (when viewed in full size, 1000 pixels wide) contains 1 million pixels, each of a different color. The human eye can distinguish about 10 million different colors.[17]
This image (when viewed in full size, 1000 pixels wide) contains 1 million pixels, each of a different color. The human eye can distinguish about 10 million different colors.[17]From the V1 blobs, color information is sent to cells in the second visual area, V2. The cells in V2 that are most strongly color tuned are clustered in the "thin stripes" that, like the blobs in V1, stain for the enzyme cytochrome oxidase (separating the thin stripes are interstripes and thick stripes, which seem to be concerned with other visual information like motion and high-resolution form). Neurons in V2 then synapse onto cells in the extended V4. This area includes not only V4, but two other areas in the posterior inferior temporal cortex, anterior to area V3, the dorsal posterior inferior temporal cortex, and posterior TEO.[18][19] (Area V4 was identified by Semir Zeki to be exclusively dedicated to color, but this has since been shown not to be the case.[20] Color processing in the extended V4 occurs in millimeter-sized color modules called globs.[18][19] This is the first part of the brain in which color is processed in terms of the full range of hues found in color space.[18][19]
Anatomical studies have shown that neurons in extended V4 provide input to the inferior temporal lobe . "IT" cortex is thought to integrate color information with shape and form, although it has been difficult to define the appropriate criteria for this claim. Despite this murkiness, it has been useful to characterize this pathway (V1 > V2 > V4 > IT) as the ventral stream or the "what pathway", distinguished from the dorsal stream ("where pathway") that is thought to analyze motion, among many other features.
Subjectivity of color perception
 Side by side comparison of the same rainbow photo in color and monochrome. The stripes of color, perceived vividly by most people, totally disappear when the color saturation is set to zero.
Side by side comparison of the same rainbow photo in color and monochrome. The stripes of color, perceived vividly by most people, totally disappear when the color saturation is set to zero.It has been established[21] that the Himba people perceive colors differently from most humans - they easily distinguish close shades of green, barely discernable for most people. The leading explanation is that the Himba created a very different color scheme which divides the spectrum to dark shades (Zuzu in Himba), very light (Vapa), Vivid blue and green (Buru) and dry colors - probably due to their specific way of life. However other explanations exist that have not been ruled out yet.
An example of the subjectivity of color occurs in a rainbow. In a rainbow (or a spectrum of light projected from a prism), the changes between wavelengths of light are smooth and continuous; there are no breaks or boundaries corresponding to the "bands of color" which are seen subjectively by the eye. A black-and-white photograph of a rainbow shows no band stucture at all, demonstrating that the number of bands, and the bands themselves, are phenomena added to nature by the eye and the brain. They are not objectively real any more than "hot" or "cold."
The cone photoreceptors, which begin the process that results in the ultimate sensation of color in the brain, are sensitive to different portions of the visible spectrum. For humans, the visible spectrum ranges approximately from 380 to 740 nm, and there are normally three types of cones. The number of colors that can be distinguished by these cones is in principle unlimited and in practice extremely large.
The sensation of color is also heavily dependent on contrast with surroundings. For example, a 'red' piece of construction paper or red filing folder does not reflect pure monochromatic "red" light.[22] Rather, it absorbs a larger fraction of other frequencies of visible light shining upon it than those in a group of frequencies that are perceived as red when they are viewed alone. However, although the red paper reflects more of light near the red end of the spectrum, it reflects some of all frequencies (not only red), and a room lit with only the reflection from a red piece of paper would soon appear to be normally lit to an adapted eye,[citation needed] and would allow visualization of all of the other colors present in the room. A red-colored paper is therefore perceived to be red mainly by contrast with its surroundings, because the human eye can distinguish between different wavelengths, and the brain creates the separate sensation of "red" as a helper in discriminating one object from another, or from its background. Color, which in nature is never constructed from pure frequencies of light, is a quality constructed by the visual brain from spectral reflectance, the objective property of objects. The advantage of color perception is the better discrimination of surfaces allowed by this aspect of visual processing, which makes quite similar surfaces into very different perceptions.
In non-human animals
The visible range and number of cone types differ between species. Mammals in general have color vision of a limited type, and are usually red-green color-blind, with only two types of cones. Humans, some primates, and some marsupials see an extended range of colors, but only by comparison with other mammals. Most non-mammalian vertebrate species distinguish different colors at least as well as humans, and many species of birds, fish, reptiles and amphibians, as well as some invertebrates, have more than three cone types and probably superior color vision to humans.
In most Catarrhini (Old World monkeys and apes — primates closely related to humans) there are three types of color receptors (known as cone cells), resulting in trichromatic color vision. These primates, like humans, are known as trichromats. Many other primates and other mammals are dichromats, which is the general color vision state for mammals that are active during the day (i.e., felines, canines, ungulates). Nocturnal mammals may have little or no color vision. Trichromat non-primate mammals are rare.[23][24]
Many invertebrates have color vision. Honey- and bumblebees have trichromatic color vision, which is insensitive to red but sensitive in ultraviolet. Papilio butterflies possess six types of photoreceptors and may have pentachromatic vision.[25] The most complex color vision system in animal kingdom has been found in stomatopods (such as the mantis shrimp) with up to 12 different spectral receptor types thought to work as multiple dichromatic units.[26]
Vertebrate animals such as tropical fish and birds sometimes have more complex color vision systems than humans; thus the many subtle colors they exhibit generally serve as direct signals between fish or between birds, and are not intended to signal mammals.[27] In the birds, tetrachromacy is achieved through up to four cone types, depending on species. Brightly colored oil droplets inside the cones shift or narrow the spectral sensitivity of the cell. It has been suggested that it is likely that pigeons are pentachromats.
Reptiles and amphibians also have four cone types (occasionally five), and probably see at least the same number of colors that humans do, or perhaps more. In addition, some nocturnal geckos have the capability of seeing color in dim light.[28]
In the evolution of mammals, segments of color vision were lost, then for a few species of primates, regained by gene duplication. Eutherian mammals other than primates (for example, dogs, cats, mammalian farm animals) generally have less-effective two-receptor (dichromatic) color perception systems, which distinguish blue, green, and yellow—but cannot distinguish oranges and reds. The adaptation to see reds is particularly important for primate mammals, since it leads to identification of fruits, and also newly sprouting reddish leaves, which are particularly nutritious.
However, even among primates, full color vision differs between New World and Old World monkeys. Old World primates, including monkeys and all apes, have vision similar to humans. New World monkeys may or may not have color sensitivity at this level: in most species, males are dichromats, and about 60% of females are trichromats, but the owl monkeys are cone monochromats, and both sexes of howler monkeys are trichromats.[29][30][31][32] Visual sensitivity differences between males and females in a single species is due to the gene for yellow-green sensitive opsin protein (which confers ability to differentiate red from green) residing on the X sex chromosome.
Several marsupials such as the fat-tailed dunnart (Sminthopsis crassicaudata) have been shown to have trichromatic color vision.[33]
Marine mammals, adapted for low-light vision, have only a single cone type and are thus monochromats.[citation needed]
Evolution
Main article: Evolution of color visionColor perception mechanisms are highly dependent on evolutionary factors, of which the most prominent is thought to be satisfactory recognition of food sources. In herbivorous primates, color perception is essential for finding proper (immature) leaves. In hummingbirds, particular flower types are often recognized by color as well. On the other hand, nocturnal mammals have less-developed color vision, since adequate light is needed for cones to function properly. There is evidence that ultraviolet light plays a part in color perception in many branches of the animal kingdom, especially insects. In general, the optical spectrum encompasses the most common electronic transitions in matter and is therefore the most useful for collecting information about the environment.
The evolution of trichromatic color vision in primates occurred as the ancestors of modern monkeys, apes, and humans switched to diurnal (daytime) activity and began consuming fruits and leaves from flowering plants.[34] Color vision, with UV discrimination, is also present in a number of arthropods – the only terrestrial animals besides the vertebrates to possess this trait.[35]
Some animals can distinguish colors in the ultraviolet spectrum. The UV spectrum falls outside the human visible range, except for some cataract surgery patients.[36] Birds, turtles, lizards, many fish and some rodents have UV receptors in their retinas.[37] These animals can see the UV patterns found on flowers and other wildlife that are otherwise invisible to the human eye.
UV and multi-dimensional vision[clarification needed] is an especially important adaptation in birds. It allows birds to spot small prey from a distance, navigate, avoid predators, and forage while flying at high speeds. Birds also utilize their broad spectrum vision to recognize other birds, and in sexual selection.[38][39]
Mathematics of color perception
A "physical color" is a combination of pure spectral colors (in the visible range). Since there are, in principle, infinitely many distinct spectral colors, the set of all physical colors may be thought of as an infinite-dimensional vector space, in fact a Hilbert space. We call this space Hcolor. More technically, the space of physical colors may be considered to be the (mathematical) cone over the simplex whose vertices are the spectral colors, with white at the centroid of the simplex, black at the apex of the cone, and the monochromatic color associated with any given vertex somewhere along the line from that vertex to the apex depending on its brightness.
An element C of Hcolor is a function from the range of visible wavelengths—considered as an interval of real numbers [Wmin,Wmax]—to the real numbers, assigning to each wavelength w in [Wmin,Wmax] its intensity C(w).
A humanly perceived color may be modeled as three numbers: the extents to which each of the 3 types of cones is stimulated. Thus a humanly perceived color may be thought of as a point in 3-dimensional Euclidean space. We call this space R3color.
Since each wavelength w stimulates each of the 3 types of cone cells to a known extent, these extents may be represented by 3 functions s(w), m(w), l(w) corresponding to the response of the S, M, and L cone cells, respectively.
Finally, since a beam of light can be composed of many different wavelengths, to determine the extent to which a physical color C in Hcolor stimulates each cone cell, we must calculate the integral (with respect to w), over the interval [Wmin,Wmax], of C(w)·s(w), of C(w)·m(w), and of C(w)·l(w). The triple of resulting numbers associates to each physical color C (which is an element in Hcolor) to a particular perceived color (which is a single point in R3color). This association is easily seen to be linear. It may also easily be seen that many different elements in the "physical" space Hcolor can all result in the same single perceived color in R3color, so a perceived color is not unique to one physical color.
Thus human color perception is determined by a specific, non-unique linear mapping from the infinite-dimensional Hilbert space Hcolor to the 3-dimensional Euclidean space R3color.
Technically, the image of the (mathematical) cone over the simplex whose vertices are the spectral colors, by this linear mapping, is also a (mathematical) cone in R3color. Moving directly away from the vertex of this cone represents maintaining the same chromaticity while increasing its intensity. Taking a cross-section of this cone yields a 2D chromaticity space. Both the 3D cone and its projection or cross-section are convex sets; that is, any mixture of spectral colors is also a color.
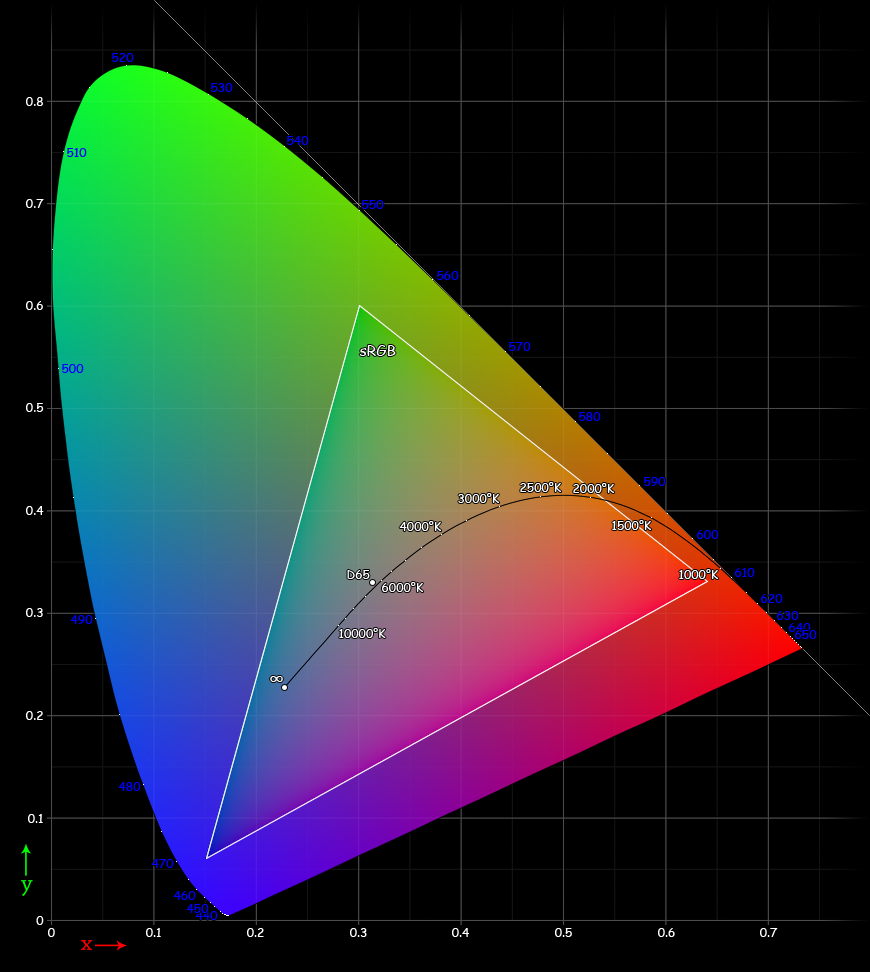
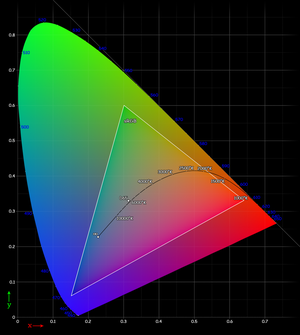 The CIE 1931 xy chromaticity diagram. The Planckian locus is shown with color temperatures labeled in degrees Kelvin. The outer curved boundary is the spectral (or monochromatic) locus, with wavelengths shown in nanometers (blue). Note that the colors in this file are being specified using sRGB. Areas outside the triangle cannot be accurately rendered because they are out of the gamut of sRGB, therefore they have been interpreted. Note that the colors depicted depend on the color space of the device you use to view the image (number of colors on your monitor, etc.), and may not be a strictly accurate representation of the color at a particular position.
The CIE 1931 xy chromaticity diagram. The Planckian locus is shown with color temperatures labeled in degrees Kelvin. The outer curved boundary is the spectral (or monochromatic) locus, with wavelengths shown in nanometers (blue). Note that the colors in this file are being specified using sRGB. Areas outside the triangle cannot be accurately rendered because they are out of the gamut of sRGB, therefore they have been interpreted. Note that the colors depicted depend on the color space of the device you use to view the image (number of colors on your monitor, etc.), and may not be a strictly accurate representation of the color at a particular position.In practice, it would be quite difficult to physiologically measure an individual's three cone responses to various physical color stimuli. Instead, a psychophysical approach is taken. Three specific benchmark test lights are typically used; let us call them S, M, and L. To calibrate human perceptual space, scientists allowed human subjects to try to match any physical color by turning dials to create specific combinations of intensities (IS, IM, IL) for the S, M, and L lights, resp., until a match was found. This needed only to be done for physical colors that are spectral (since a linear combination of spectral colors will be matched by the same linear combination of their (IS, IM, IL) matches. Note that in practice, often at least one of S, M, L would have to be added with some intensity to the physical test color, and that combination matched by a linear combination of the remaining 2 lights. Across different individuals (without color blindness), the matchings turned out to be nearly identical.
By considering all the resulting combinations of intensities (IS, IM, IL) as a subset of 3-space, a model for human perceptual color space is formed. (Note that when one of S, M, L had to be added to the test color, its intensity was counted as negative.) Again, this turns out to be a (mathematical) cone, not a quadric, but rather all rays through the origin in 3-space passing through a certain convex set. Again, this cone has the property that moving directly away from the origin corresponds to increasing the intensity of the S, M, L lights proportionately. Again, a cross-section of this cone is a planar shape that is (by definition) the space of "chromaticities" (informally: distinct colors); one particular such cross section, corresponding to constant X+Y+Z of the CIE 1931 color space, gives the CIE chromaticity diagram.
It should be noted that this system implies that for any hue or non-spectral color not on the boundary of the chromaticity diagram, there are infinitely many distinct physical spectra that are all perceived as that hue or color. So, in general there is no such thing as the combination of spectral colors that we perceive as (say) a specific version of tan; instead there are infinitely many possibilities that produce that exact color. The boundary colors that are pure spectral colors can be perceived only in response to light that is purely at the associated wavelength, while the boundary colors on the "line of purples" can each only be generated by a specific ratio of the pure violet and the pure red at the ends of the visible spectral colors.
The CIE chromaticity diagram is horseshoe-shaped, with its curved edge corresponding to all spectral colors (the spectral locus), and the remaining straight edge corresponding to the most saturated purples, mixtures of red and violet.
Chromatic adaptation
Main article: Chromatic adaptationIn color science, chromatic adaptation is the estimation of the representation of an object under a different light source than the one in which it was recorded. A common application is to find a chromatic adaptation transform (CAT) that will make the recording of a neutral object appear neutral (color balance), while keeping other colors also looking realistic.[40] For example, chromatic adaptation transforms are used when converting images between ICC profiles with different white points. Adobe Photoshop, for example, uses the Bradford CAT.[41]
In color vision, chromatic adaptation refers to color constancy; the ability of the visual system to preserve the appearance of an object under a wide range of light sources.[42]
See also
References
- ^ "Eye, human." Encyclopædia Britannica 2006 Ultimate Reference Suite DVD, 2009.
- ^ Neitz J, Jacobs GH (1986). "Polymorphism of the long-wavelength cone in normal human color vision". Nature 323 (6089): 623–5. doi:10.1038/323623a0. PMID 3773989. http://www.nature.com/nature/journal/v323/n6089/abs/323623a0.html.
- ^ Jacobs GH (January 1996). "Primate photopigments and primate color vision". Proc. Natl. Acad. Sci. U.S.A. 93 (2): 577–81. doi:10.1073/pnas.93.2.577. PMC 40094. PMID 8570598. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=40094.
- ^ Hering, Ewald (1872). "Zur Lehre vom Lichtsinne". Sitzungsberichte der Mathematisch–Naturwissenschaftliche Classe der Kaiserlichen Akademie der Wissenschaften (K.-K. Hof- und Staatsdruckerei in Commission bei C. Gerold's Sohn) LXVI. Band (III Abtheilung). http://books.google.com/?id=u5MCAAAAYAAJ&pg=PA5&lpg=PA5&dq=1872+hering+ewald+Zur+Lehre+vom+Lichtsinne.+Sitzungsberichte+der+kaiserlichen+Akademie+der+Wissenschaften.+Mathematisch%E2%80%93naturwissenschaftliche+Classe,.
- ^ Ali, M.A. & Klyne, M.A. (1985), p.168
- ^ Wyszecki, Günther; Stiles, W.S. (1982). Color Science: Concepts and Methods, Quantitative Data and Formulae (2nd ed.). New York: Wiley Series in Pure and Applied Optics. ISBN 0-471-02106-7.
- ^ R. W. G. Hunt (2004). The Reproduction of Colour (6th ed.). Chichester UK: Wiley–IS&T Series in Imaging Science and Technology. pp. 11–2. ISBN 0-470-02425-9.
- ^ Verrelli BC, Tishkoff SA (September 2004). "Signatures of Selection and Gene Conversion Associated with Human Color Vision Variation". Am. J. Hum. Genet. 75 (3): 363–75. doi:10.1086/423287. PMC 1182016. PMID 15252758. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1182016.
- ^ Roth, Mark (2006). "Some women may see 100 million colors, thanks to their genes" Post-Gazette.com
- ^ R.W. Rodieck, "The First Steps in Seeing". Sinauer Associates, Inc., Sunderland, Massachusetts, USA, 1998.
- ^ SH Hendry, RC Reid, "The Koniocellular Pathway in Primate Vision". Annual Reviews Neuroscience, 2000, vol. 23, pp. 127-53
- ^ Nigel W. Daw (17 November 1967). "Goldfish Retina: Organization for Simultaneous Color Contrast". Science 158 (3803): 942–4. doi:10.1126/science.158.3803.942. PMID 6054169.
- ^ Bevil R. Conway (2002). Neural Mechanisms of Color Vision: Double-Opponent Cells in the Visual Cortex. Springer. ISBN 1402070926. http://books.google.com/?id=pFodUlHfQmcC&pg=PR7&dq=goldfish+retina+by+Nigel-Daw.
- ^ Conway BR (15 April 2001). "Spatial structure of cone inputs to color cells in alert macaque primary visual cortex (V-1)". J. Neurosci. 21 (8): 2768–83. PMID 11306629. http://www.jneurosci.org/content/21/8/2768.full.
- ^ John E. Dowling (2001). Neurons and Networks: An Introduction to Behavioral Neuroscience. Harvard University Press. ISBN 0674004620. http://books.google.com/?id=adeUwgfwdKwC&pg=PA376&dq=Margaret+Livingstone+David+Hubel+double+opponent+blobs.
- ^ McCann, M., ed. 1993. Edwin H. Land's Essays. Springfield, Va.: Society for Imaging Science and Technology.
- ^ Judd, Deane B.; Wyszecki, Günter (1975). Color in Business, Science and Industry. Wiley Series in Pure and Applied Optics (3rd ed.). New York: Wiley-Interscience. p. 388. ISBN 0471452122.
- ^ a b c Conway BR, Moeller S, Tsao DY (2007). "Specialized color modules in macaque extrastriate cortex". Neuron 56 (3): 560–73. doi:10.1016/j.neuron.2007.10.008. PMID 17988638.
- ^ a b c Conway BR, Tsao DY (2009). "Color-tuned neurons are spatially clustered according to color preference within alert macaque posterior inferior temporal cortex". Proc Natl Acad Sci U S A 106 (42): 18035–18039. doi:10.1073/pnas.0810943106. PMC 2764907. PMID 19805195. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2764907.
- ^ John Allman and Steven W. Zucker (1993). "On cytochrome oxidase blobs in visual cortex". In Laurence Harris and Michael Jenkin, editors. Spatial Vision in Humans and Robots: The Proceedings of the 1991 York Conference. Cambridge University Press. ISBN 0521430712. http://books.google.com/?id=eWBiKaOCNIYC&pg=PA34&dq=v4+zeki+color.
- ^ Roberson, Davidoff, Davies & Shapiro. referred by Debi Roberson, University of Essex 2011
- ^ Wright, W. D. (1967). The rays are not coloured: essays on the science and vision and colour. Bristol: Hilger. ISBN 0-85274-068-9.
- ^ Ali, Mohamed Ather; Klyne, M.A. (1985). Vision in Vertebrates. New York: Plenum Press. pp. 174–175. ISBN 0-306-42065-1.
- ^ For .pdf online review see: [1] THE DISTRIBUTION AND NATURE OF COLOUR VISION AMONG THE MAMMALS. GERALD H. JACOBS Biological Reviews Volume 68, Issue 3, pages 413–471, August 1993. DOI: 10.1111/j.1469-185X.1993.tb00738.x
- ^ Arikawa K (November 2003). "Spectral organization of the eye of a butterfly, Papilio". J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 189 (11): 791–800. doi:10.1007/s00359-003-0454-7. PMID 14520495. http://www.springerlink.com/content/whjepqnhpulyeevk/.
- ^ Cronin TW, Marshall NJ (1989). "A retina with at least ten spectral types of photoreceptors in a mantis shrimp". Nature 339 (6220): 137–40. doi:10.1038/339137a0. http://www.nature.com/nature/journal/v339/n6220/abs/339137a0.html.
- ^ Kelber A, Vorobyev M, Osorio D (February 2003). "Animal color vision—behavioural tests and physiological concepts". Biol Rev Camb Philos Soc 78 (1): 81–118. doi:10.1017/S1464793102005985. PMID 12620062.
- ^ Roth, Lina S. V.; Lundström, Linda; Kelber, Almut; Kröger, Ronald H. H.; Unsbo, Peter (March 30, 2009). "The pupils and optical systems of gecko eyes". Journal of Vision 9 (3:27): 1–11. doi:10.1167/9.3.27. PMID 19757966. http://www.journalofvision.org/content/9/3/27.
- ^ Jacobs G. H., Deegan J. F. (2001). "Photopigments and color vision in New World monkeys from the family Atelidae". Proceedings of the Royal Society of London, Series B 268 (1468): 695–702. doi:10.1098/rspb.2000.1421.
- ^ Jacobs G. H., Deegan J. F., Neitz , Neitz J., Crognale M. A. (1993). "Photopigments and color vision in the nocturnal monkey, Aotus". Vision Research 33 (13): 1773–1783. doi:10.1016/0042-6989(93)90168-V. PMID 8266633.
- ^ Mollon J. D., Bowmaker J. K., Jacobs G. H. (1984). "Variations of color vision in a New World primate can be explained by polymorphism of retinal photopigments". Proceedings of the Royal Society of London, Series B 222 (1228): 373–399. doi:10.1098/rspb.1984.0071.
- ^ Sternberg, Robert J. (2006): Cognitive Psychology. 4th Ed. Thomson Wadsworth.
- ^ Arrese CA, Beazley LD, Neumeyer C (March 2006). "Behavioural evidence for marsupial trichromacy". Curr. Biol. 16 (6): R193–4. doi:10.1016/j.cub.2006.02.036. PMID 16546067.
- ^ Pinker, Steven (1997). How the Mind Works. New York: Norton. p. 191. ISBN 0-393-04535-8.
- ^ Koyanagi, M.; Nagata, T.; Katoh, K.; Yamashita, S.; Tokunaga, F. (2008). "Molecular Evolution of Arthropod Color Vision Deduced from Multiple Opsin Genes of Jumping Spiders". Journal of Molecular Evolution 66 (2): 130. doi:10.1007/s00239-008-9065-9. PMID 18217181.
- ^ David Hambling (May 30, 2002). "Let the light shine in: You don't have to come from another planet to see ultraviolet light". EducationGuardian.co.uk. http://www.guardian.co.uk/science/2002/may/30/medicalscience.research.
- ^ Jacobs GH, Neitz J, Deegan JF (1991). "Retinal receptors in rodents maximally sensitive to ultraviolet light". Nature 353 (6345): 655–6. doi:10.1038/353655a0. PMID 1922382. http://www.nature.com/nature/journal/v353/n6345/abs/353655a0.html.
- ^ FJ Varela, AG Palacios, and TM Goldsmith (1993). Bischof, Hans-Joachim; Zeigler, H. Philip. ed. Vision, brain, and behavior in birds. Cambridge, Mass: MIT Press. pp. 77–94. ISBN 0-262-24036-X.
- ^ IC Cuthill, JC Partridge, ATD Bennett, SC Church, NS Hart, and S Hunt (2000). "Ultraviolet Vision in Birds". Advances in the Study of Behavior. 29. pp. 159–214.
- ^ Süsstrunk, Sabine. Chromatic Adaptation
- ^ Lindbloom, Bruce. Chromatic Adaptation
- ^ Fairchild, Mark D. (2005). "8. Chromatic Adaptation". Color Appearance Models. Wiley. p. 146. ISBN 0470012161. http://books.google.com/?id=8_TxzK2B-5MC&pg=PA146&dq=%22chromatic+adaptation%22.
External links
- Peter Gouras, "Color Vision", Webvision, University of Utah School of Medicine, May 2009.
- Kenneth R. Koehler, "Spectral Sensitivity of the Eye", College Physics for Students of Biology and Chemistry, University of Cincinnati Raymond Walters College, 1996.
- James T. Fulton, "The Human is a Blocked Tetrachromat", Neural Concepts, July 2009.
- Vurdlak, "Mega Color Blindness Test", Mighty Optical Illusions, March 2009.
- Clive Maxfield and Alvin Brown, "Color Vision: One of Nature's Wonders", DIYCalculator.com, 2006.
- Chenguang Lu, "The decoding model: a symmetrical model of color vision".
- Egopont, "Color Vision Test".
Physiology: Eye and ear physiology Vision Gaze · Accommodation · Intraocular pressure · Visual field
Color vision · Color blindness · Opponent process
Monochromacy · Dichromacy · Trichromacy · Tetrachromacy · PentachromacyAuditory system M: EYE
anat(g/a/p)/phys/devp/prot
noco/cong/tumr, epon
proc, drug(S1A/1E/1F/1L)
M: EAR
anat(e/p)/phys/devp
noco/cong, epon
proc, drug(S2)
Color topics Color perception Color vision · Color blindness · Visible spectrum · Color constancy · Color term · Color theory · Complementary color
Color space Basic colors Related Mental processes Cognition Perception Amodal perception · Color perception · Depth perception · Visual perception · Form perception · Haptic perception · Speech perception · Perception as Interpretation · Numeric Value of Perception · Pitch perception · Harmonic perception · Social perceptionMemory Other Categories:







Wikimedia Foundation. 2010.