- Orchidaceae
-
"Orchid" redirects here. For other uses, see Orchid (disambiguation).
Orchidaceae
Temporal range: 80 Ma Late Cretaceous - Recent
Color plate from Ernst Haeckel's Kunstformen der Natur Scientific classification Kingdom: Plantae (unranked): Angiosperms (unranked): Monocots Order: Asparagales Family: Orchidaceae
Juss.Type genus Orchis
Tourn. ex L.Subfamilies - Apostasioideae Horaninov
- Cypripedioideae Kosteletzky
- Epidendroideae Kosteletzky
- Orchidoideae Eaton
- Vanilloideae Szlachetko
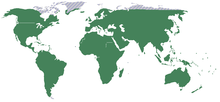
Distribution range of family Orchidaceae The Orchidaceae, commonly referred to as the orchid family, is a morphologically diverse and widespread family of monocots in the order Asparagales. Along with the Asteraceae, it is one of the two largest families of flowering plants, with between 21,950 and 26,049 currently accepted species, found in 880 genera.[1][2] Selecting which of the two families is larger remains elusive because of the difficulties associated with putting hard species numbers on such enormous groups. Regardless, the number of orchid species equals more than twice the number of bird species, and about four times the number of mammal species. It also encompasses about 6–11% of all seed plants.[3] The largest genera are Bulbophyllum (2,000 species), Epidendrum (1,500 species), Dendrobium (1,400 species) and Pleurothallis (1,000 species).
The family also includes Vanilla (the genus of the vanilla plant), Orchis (type genus), and many commonly cultivated plants such as Phalaenopsis and Cattleya. Moreover, since the introduction of tropical species in the 19th century, horticulturists have produced more than 100,000 hybrids and cultivars.
Contents
Etymology
The name comes from the Greek ὄρχις (órkhis), literally meaning "testicle", because of the shape of the root.[4][5]. Linnaeus categorized the family as Orchidaceae. Orchid was introduced in 1845 by John Lindley in School Botany, due to an incorrect attempt to extract the Latin stem (orchis) from Orchidaceae.
The Greek myth of Orchis explains the origin of the plants. Orchis, the son of a nymph and a satyr, came upon a festival of Dionysios (Bacchus) in the forest. He drank too much, and attempted to rape a priestess of Dionysios. For his insult, he was torn apart by the Bacchanalians. His father prayed for him to be restored, but the gods instead changed him into a flower.
These flowers were previously called Orchis, Satyrion (Satyrion feminina), or "ballockwort".
Distribution
Orchidaceae are cosmopolitan, occurring in almost every habitat apart from glaciers. The world's richest concentration of orchid varieties is found in the Himalayan region of Nepal. The great majority are to be found in the tropics, mostly Asia, South America and Central America, but they are also found above the Arctic Circle, in southern Patagonia, and even two species of Nematoceras on Macquarie Island, close to Antarctica.
The following list gives a rough overview of their distribution:
- tropical Asia: 260 to 300 genera
- tropical America: 250 to 270 genera
- tropical Africa: 230 to 270 genera
- Oceania: 50 to 70 genera
- Europe and temperate Asia: 40 to 60 genera
- North America: 20 to 26 genera
Taxonomy
Main article: Taxonomy of the Orchid familyThe taxonomy of this family is in constant flux, as new studies continue to identify more classificatory elements. The Orchidaceae is currently placed in the order Asparagales by the APG III system of 2009.
Five subfamilies are recognised. The cladogram has been made according to the APG system:
Apostasioideae: 2 genera and 16 species, south-western Asia
Cypripedioideae: 5 genera and 130 species, from the temperate regions of the world, as well as tropical America and tropical Asia
Monandrae Vanilloideae: 15 genera and 180 species, humid tropical and subtropical regions, eastern North America
Epidendroideae: more than 500 genera and more or less 20,000 species, cosmopolitan
Orchidoideae: 208 genera and 3,630 species, cosmopolitan
Characteristics
Orchids are easily distinguished from other plants, as they share some very evident apomorphies. Among these are: bilateral symmetry (zygomorphism), many resupinate flowers, a nearly always highly modified petal (labellum), fused stamens and carpels, and extremely small seeds.
Stem and Roots
All orchids are perennial herbs, lack any permanent woody structure, and can grow according to two patterns:
- Monopodial: The stem grows from a single bud, leaves are added from the apex each year and the stem grows longer accordingly. The stem of orchids with a monopodial growth can reach several metres in length, as in Vanda and Vanilla.
- Sympodial: The plant produces a series of adjacent shoots which grow to a certain size, bloom and then stop growing, to be then replaced. Sympodial orchids grow laterally rather than vertically, following the surface of their support. The growth continues by development of new leads, with their own leaves and roots, sprouting from or next to those of the previous year, as in Cattleya. While a new lead is developing, the rhizome may start its growth again from a so-called 'eye', an undeveloped bud, thereby branching.
 Anacamptis lactea showing the two tubers
Anacamptis lactea showing the two tubers
Terrestrial orchids may be rhizomatous or form corms or tubers. The root caps of terrestrials are smooth and white.
Some sympodial terrestrials, such as Orchis and Ophrys, have two subterranean tuberous roots. One is used as a food reserve for wintry periods, and provides for the development of the other one, from which visible growth develops.
In warm and humid climates, many terrestrial orchids do not need pseudobulbs.
Epiphytic orchids have modified aerial roots that can sometimes be a few meters long. In the older parts of the roots, a modified spongy epidermis, called velamen, has the function to absorb humidity. It is made of dead cells and can have a silvery-grey, white or brown appearance. In some orchids, the velamen includes spongy and fibrous bodies near the passage cells, called tilosomes.
The cells of the root epidermis grow at a right angle to the axis of the root to allow them to get a firm grasp on their support. Nutrients mainly come from animal droppings and other organic detritus on their supporting surfaces.
 The pseudobulb of Prosthechea fragrans
The pseudobulb of Prosthechea fragransThe base of the stem of sympodial epiphytes, or in some species essentially the entire stem, may be thickened to form a pseudobulb that contains nutrients and water for drier periods.
The pseudobulb has a smooth surface with lengthwise grooves, and can have different shapes, often conical or oblong. Its size is very variable; in some small species of Bulbophyllum, it is no longer than two millimeters, while in the largest orchid in the world, Grammatophyllum speciosum (giant orchid), it can reach three meters. Some Dendrobium species have long, canelike pseudobulbs with short, rounded leaves over the whole length; some other orchids have hidden or extremely small pseudobulbs, completely included inside the leaves.
With ageing, the pseudobulb sheds its leaves and becomes dormant. At this stage it is often called a backbulb. A pseudobulb then takes over, exploiting the last reserves accumulated in the backbulb, which eventually dies off, too. A pseudobulb typically lives for about five years.
Leaves
 A close-up of a Phalaenopsis orchid leaf, the parallel veins and cuticle are visible.
A close-up of a Phalaenopsis orchid leaf, the parallel veins and cuticle are visible.Like most monocots, orchids generally have simple leaves with parallel veins, although some Vanilloideae have a reticulate venation. Leaves may be ovate, lanceolate, or orbiculate, and very variable in size. Their characteristics are often diagnostic. They are normally alternate on the stem, often plicate, and have no stipules. Orchid leaves often have siliceous bodies called stegmata in the vascular bundle sheaths (not present in the Orchidoideae) and are fibrous.
The structure of the leaves corresponds to the specific habitat of the plant. Species that typically bask in sunlight, or grow on sites which can be occasionally very dry, have thick, leathery leaves and the laminae are covered by a waxy cuticle to retain their necessary water supply. Shade species, on the other hand, have long, thin leaves.
The leaves of most orchids are perennial, that is, they live for several years, while others, especially those with plicate leaves, shed them annually and develop new leaves together with new pseudobulbs, as in Catasetum.
The leaves of some orchids are considered ornamental. The leaves of the Macodes sanderiana, a semiterrestrial or lithophyte, show a sparkling silver and gold veining on a light green background. The cordate leaves of Psychopsiella limminghei are light brownish-green with maroon-puce markings, created by flower pigments. The attractive mottle of the leaves of lady's slippers from tropical and subtropical Asia (Paphiopedilum), is caused by uneven distribution of chlorophyll. Also, Phalaenopsis schilleriana is a pastel pink orchid with leaves spotted dark green and light green. The jewel orchid (Ludisia discolor) is grown more for its colorful leaves than its white flowers.
Some orchids, as Dendrophylax lindenii (ghost orchid), Aphyllorchis and Taeniophyllum depend on their green roots for photosynthesis and lack normally developed leaves, as do all of the heterotrophic species.
Orchids of the genus Corallorhiza (coralroot orchids) lack leaves altogether and instead wrap their roots around the roots of mature trees and use specialized fungi to harvest sugars.[6]
Flowers
 Dactylorhiza sambucina, Orchidoideae for reference
Dactylorhiza sambucina, Orchidoideae for referenceOrchidaceae are well known for the many structural variations in their flowers.
Some orchids have single flowers, but most have a racemose inflorescence, sometimes with a large number of flowers. The flowering stem can be basal, that is, produced from the base of the tuber, like in Cymbidium, apical, meaning it grows from the apex of the main stem, like in Cattleya, or axillary, from the leaf axil, as in Vanda.
As an apomorphy of the clade, orchid flowers are primitively zygomorphic (bilaterally symmetrical), although in some genera like Mormodes, Ludisia and Macodes, this kind of symmetry may be difficult to notice.
The orchid flower, like most flowers of monocots, has two whorls of sterile elements. The outer whorl has three sepals and the inner whorl has three petals. The sepals are usually very similar to the petals (and thus called tepals, 1), but may be completely distinct.
The upper medial petal, called the labellum or lip (6), is always modified and enlarged. The inferior ovary (7) or the pedicel usually rotates 180 degrees, so that the labellum, goes on the lower part of the flower, thus becoming suitable to form a platform for pollinators. This characteristic, called resupination, occurs primitively in the family and is considered apomorphic (the torsion of the ovary is very evident from the picture). Some orchids have secondarily lost this resupination, e. g. Zygopetalum and Epidendrum secundum.
The normal form of the sepals can be found in Cattleya, where they form a triangle. In Paphiopedilum (Venus slippers), the lower two sepals are fused into a synsepal, while the lip has taken the form of a slipper. In Masdevallia, all the sepals are fused.
Orchid flowers with abnormal numbers of petals or lips are called peloric. Peloria is a genetic trait, but its expression is environmentally influenced and may appear random.
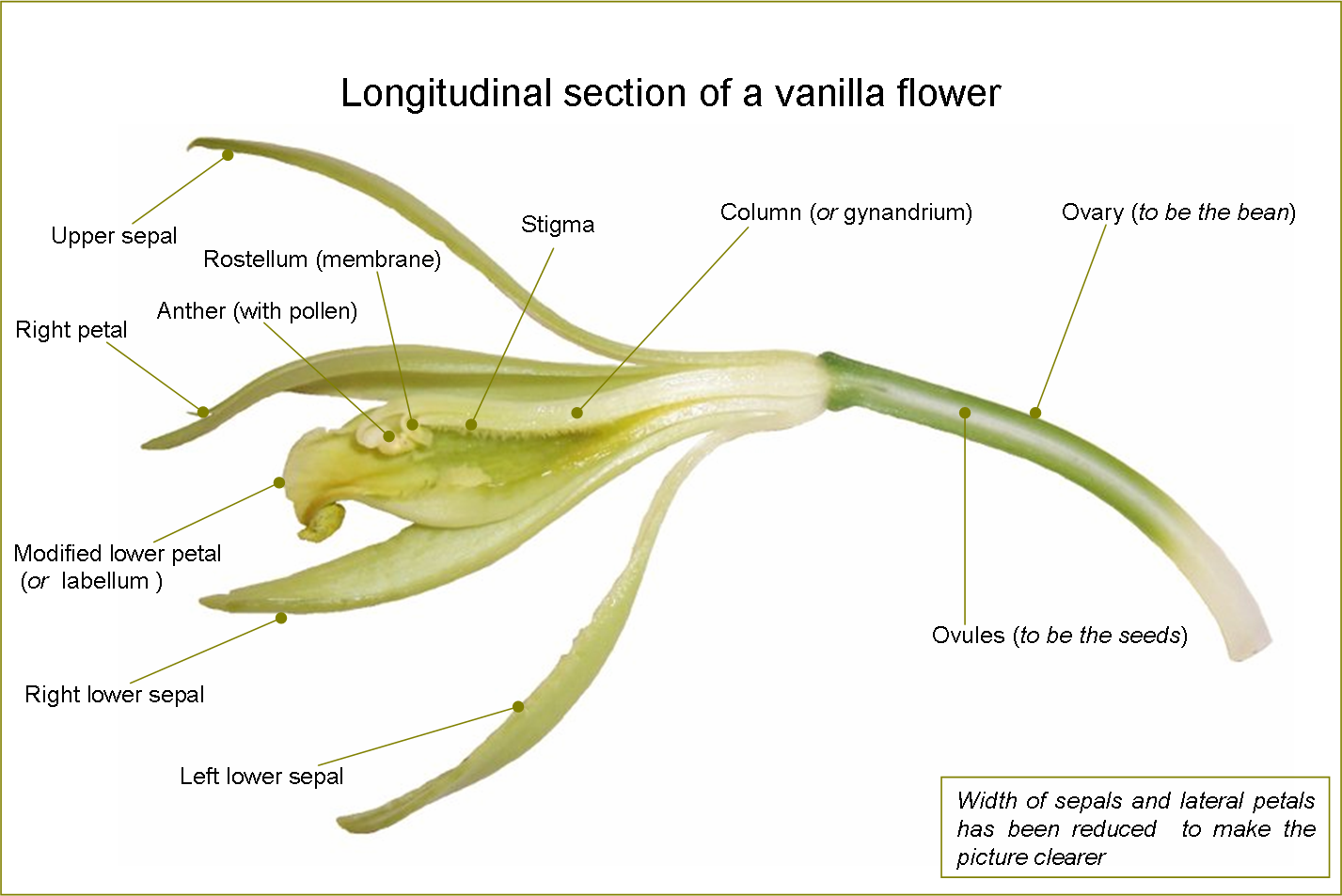
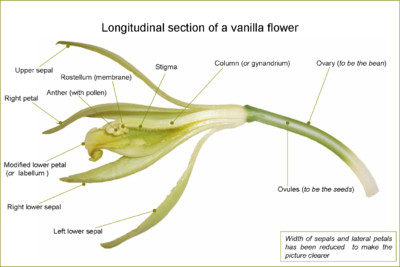 Longitudinal section of a flower of Vanilla planifolia
Longitudinal section of a flower of Vanilla planifoliaOrchid flowers primitively had three stamens, but this situation is now limited to the genus Neuwiedia. Apostasia and the Cypripedioideae have two stamens, the central one being sterile and reduced to a staminode. All of the other orchids, the clade called Monandria, retain only the central stamen, the others being reduced to staminodes (4). The filaments of the stamens are always adnate (fused) to the style to form cylindrical structure called the gynostemium or column (2). In the primitive Apostasioideae, this fusion is only partial; in the Vanilloideae, it is more deep; in Orchidoideae and Epidendroideae, it is total. The stigma (9) is very asymmetrical, as all of its lobes are bent towards the centre of the flower and lay on the bottom of the column.
Pollen is released as single grains, like in most other plants, in the Apostasioideae, Cypripedioideae and Vanilloideae. In the other subfamilies, that comprise the great majority of orchids, the anther (3), carries and two pollinia.
A pollinium is a waxy mass of pollen grains held together by the glue-like alkaloid viscin, containing both cellulosic strands and mucopolysaccharides. Each pollinium is connected to a filament which can take the form of a caudicle, as in Dactylorhiza or Habenaria, or a stipe, as in Vanda. Caudicles or stipes hold the pollinia to the viscidium, a sticky pad which sticks the pollinia to the body of pollinators.
At the upper edge of the stigma of single-anthered orchids, in front of the anther cap, there is the rostellum (5), a slender extension involved in the complex pollination mechanism.
As aforementioned, the ovary is always inferior (located behind the flower). It is three-carpelate and one or, more rarely, three-partitioned, with parietal placentation (axile in the Apostasioideae).
Fruits and Seeds
 Cross-section of an orchid capsule, the longitudinal slits
Cross-section of an orchid capsule, the longitudinal slitsThe ovary typically develops into a capsule that is dehiscent by three or six longitudinal slits, while remaining closed at both ends. The ripening of a capsule can take two to 18 months.
The seeds are generally almost microscopic and very numerous, in some species over a million per capsule. After ripening, they blow off like dust particles or spores. They lack endosperm and must enter symbiotic relationships with various mycorrhizal basidiomyceteous fungi that provide them the necessary nutrients to germinate, so that all orchid species are mycoheterotrophic during germination and reliant upon fungi to complete their lifecycles.
 Closeup of a Phalaenopsis blossom
Closeup of a Phalaenopsis blossomAs the chance for a seed to meet a fitting fungus is very small, only a minute fraction of all the seeds released grow into adult plants. In cultivation, germination typically takes weeks, while there is a report of one paphiopedilum that took fifteen years.
Horticultural techniques have been devised for germinating seeds on a nutrient-containing gel, eliminating the requirement of the fungus for germination, greatly aiding the propagation of ornamental orchids.
The main component for the sowing of orchids in artificial conditions is the agar agar. The substance is put together with some type of carbohydrate (actually, some kind of glucose) which provides qualitative organic feed. Such substance may be banana, pineapple, peach or even tomato puree or coconut milk. After the "cooking" of the agar agar (it has to be cooked in sterile conditions), the mix is poured into test tubes or jars where the substance begins to gel.
Reproduction
Pollination
The complex mechanisms which orchids have evolved to achieve cross-pollination were investigated by Charles Darwin and described in his 1862 book Fertilisation of Orchids. Orchids have developed highly specialized pollination systems, thus the chances of being pollinated are often scarce, so orchid flowers usually remain receptive for very long periods, and most orchids deliver pollen in a single mass. Each time pollination succeeds, thousands of ovules can be fertilized.
Pollinators are often visually attracted by the shape and colours of the labellum. The flowers may produce attractive odours. Although absent in most species, nectar may be produced in a spur (8) of the labellum, on the point of the sepals or in the septa of the ovary, the most typical position amongst the Asparagales.
In orchids that produce pollinia, pollination happens as some variant of the following. When the pollinator enters into the flower, it touches a viscidium, which promptly sticks to its body, generally on the head or abdomen. While leaving the flower, it pulls the pollinium out of the anther, as it is connected to the viscidium by the caudicle or stipe. The caudicle then bends and the pollinium is moved forwards and downwards. When the pollinator enters another flower of the same species, the pollinium has taken such position that it will stick to the stigma of the second flower, just below the rostellum, pollinating it. The possessors of orchids may be able to reproduce the process with a pencil, small paintbrush, or other similar device.
 Ophrys apifera is about to self-pollinate
Ophrys apifera is about to self-pollinateSome orchids mainly or totally rely on self-pollination, especially in colder regions where pollinators are particularly rare. The caudicles may dry up if the flower has not been visited by any pollinator, and the pollinia then fall directly on the stigma. Otherwise, the anther may rotate and then enter the stigma cavity of the flower (as in Holcoglossum amesianum).
The labellum of the Cypripedioideae is poke-shaped, and has the function to trap visiting insects. The only exit leads to the anthers that deposit pollen on the visitor.
In some extremely specialized orchids, such as the Eurasian genus Ophrys, the labellum is adapted to have a colour, shape and odour which attracts male insects via mimicry of a receptive female. Pollination happens as the insect attempts to mate with flowers.
Many neotropical orchids are pollinated by male orchid bees, which visit the flowers to gather volatile chemicals they require to synthesize pheromonal attractants. Each type of orchid places the pollinia on a different body part of a different species of bee, so as to enforce proper cross-pollination.
An underground orchid in Australia, Rhizanthella slateri, is never exposed to light, and depends on ants and other terrestrial insects to pollinate it.
Catasetum, a genus discussed briefly by Darwin, actually launches its viscid pollinia with explosive force when an insect touches a seta, knocking the pollinator off the flower.
After pollination, the sepals and petals fade and wilt, but they usually remain attached to the ovary.
Asexual Reproduction
Some species, such as Phalaenopsis, Dendrobium and Vanda, produce offshoots or plantlets formed from one of the nodes along the stem, through the accumulation of growth hormones at that point. These shoots are known as keiki.
Evolution
A study in the scientific journal Nature [7] has hypothesized that the origin of orchids goes back much longer than originally expected. An extinct species of stingless bee, Proplebeia dominicana, was found trapped in Miocene amber from about 15-20 million years ago. The bee was carrying pollen of a previously unknown orchid taxon, Meliorchis caribea, on its wings. This find is the first evidence of fossilised orchids to date.[7] The extinct orchid M. caribea has been placed within the extant tribe Cranichideae, subtribe Goodyerinae (subfamily Orchidoideae).
This indicates orchids may have arisen 76 to 84 million years ago during the Late Cretaceous. In other words, they may have coexisted with dinosaurs. It also shows insects were active pollinators of orchids then. According to Chase et al. (2001), the overall biogeography and phylogenetic patterns of Orchidaceae show they are even older and may go back roughly 100 million years.[8]
Using the molecular clock method, it was possible to determine the age of the major branches of the orchid family. This also confirmed that the subfamily Vanilloideae is a branch at the basal dichotomy of the monandrous orchids, and must have evolved very early in the evolution of the family. Since this genus occurs worldwide in tropical and subtropical regions, from tropical America to tropical Asia, New Guinea and West Africa, and the continents began to split about 100 million years ago, significant biotic exchange must have occurred after this split (since the age of Vanilla is estimated at 60 to 70 million years).
Ecology
A majority of orchids are perennial epiphytes, which grow anchored to trees or shrubs in the tropics and subtropics. Species such as Angraecum sororium are lithophytes[9], growing on rocks or very rocky soil. Other orchids (including the majority of temperate Orchidaceae) are terrestrial and can be found in habitat areas such as grasslands or forest.
Some orchids, such as Neottia and Corallorhiza, lack chlorophyll, so are unable to photosynthesize. Instead, these species obtain energy and nutrients by parasitising soil fungi through the formation of orchid mycorrhizas. The fungi involved include those that form ectomycorrhizas with trees and other woody plants, parasites such as Armillaria, and saprotrophs.[10] These orchids are known as myco-heterotrophs, but were formerly (incorrectly) described as saprophytes due to the belief that they gained their nutrition by breaking down organic matter. While only a few species are achlorophyllous holoparasites, all orchids are myco-heterotrophic during germination and seedling growth, and even photosynthetic adult plants may continue to obtain carbon from their mycorrhizal fungi.
Uses
Further information: Vanilla Vanilla fruits drying
Vanilla fruits dryingThe dried seed pods of one orchid genus, Vanilla, are commercially important as flavoring in baking, for perfume manufacture and aromatherapy.
The underground tubers of terrestrial orchids [mainly Orchis mascula (early purple orchid)] are ground to a powder and used for cooking, such as in the hot beverage salep or in the Turkish frozen treat dondurma. The name salep has been claimed to come from the Arabic expression ḥasyu al-tha`lab, "fox testicles", but it appears more likely the name comes directly from the Arabic name saḥlab. The similarity in appearance to testes naturally accounts for salep being considered an aphrodisiac.
The dried leaves of Jumellea fragrans are used to flavor rum on Reunion Island.
The scent of orchids is frequently analysed by perfumers (using headspace technology and gas-liquid chromatography) to identify potential fragrance chemicals.
The other important use of orchids is their cultivation for the enjoyment of the flowers. Most cultivated orchids are tropical or subtropical, but quite a few which grow in colder climates can be found on the market. Temperate species available at nurseries include Ophrys apifera (bee orchid), Gymnadenia conopsea (fragrant orchid), Anacamptis pyramidalis (pyramidal orchid) and Dactylorhiza fuchsii (common spotted orchid).
Orchids of all types have also often been sought by collectors of both species and hybrids. As such, many hundreds of societies and clubs worldwide have been established. These can be small, local clubs such as the Sutherland Shire Orchid Society, or larger, national organisations such as the American Orchid Society. Both serve to encourage cultivation and collection of orchids, but some go further by concentrating on conservation or research.
The term "botanical orchid" loosely denotes those small-flowered, tropical orchids belonging to several genera (not necessarily related to each other) that do not fit into the "florist" orchid category. A few of these genera contain enormous numbers of species. Some, such as Pleurothallis and Bulbophyllum, contain approximately 1700 and 2000 species, respectively, and are often extremely vegetatively diverse. The primary use of the term is among orchid hobbyists wishing to describe unusual species they grow, though it is also used to distinguish naturally occurring orchid species from horticulturally created hybrids.
Traditional Medicinal Uses
Orchids have been used in traditional medicine in an effort to treat many diseases and ailments. They have been used as a source of herbal remedies in China since 2800 BC. Gastrodia elata is one of the three orchids listed in the earliest known Chinese Materia Medica (Shennon bencaojing) (c. 100 AD). Theophrastus mentions orchids in his Enquiry into Plants (372–286 BC).
In the recent years, a number of studies have been published on anticancer activity of the chemical moscatilin, which is found in the stems of the orchid species Dendrobrium.[11]
Symbolism
Orchids have many associations with symbolic values. For example, the orchid is the City Flower of Shaoxing, China.
In Chinese culture, the bamboo, plum blossom, orchid, and chrysanthemum (often known as méi lán zhú jú 梅兰竹菊) are collectively referred to as the Four Gentlemen. These four plants also represent the four seasons and, in Confucian ideology, four aspects of the junzi ("prince" or "noble one").
Genera
Main article: List of Orchidaceae generaThe following are amongst the most notable genera of the orchid family:
- Ascocenda
- Ascocentrum
- Ascoglossum
- Australorchis
- Auxopus
- Baptistonia
- Barbrodia
- Barkeria
- Barlia
- Bartholina
- Beloglottis
- Biermannia
- Bletilla
- Brassavola
- Brassia
- Bulbophyllum
- Calypso
- Catasetum
- Cattleya
- Cirrhopetalum
- Cleisostoma
- Clowesia
- Coelogyne
- Coryanthes
- Cymbidium
- Cyrtopodium
- Cypripedium
- Dactylorhiza
- Dendrobium
- Disa
- Dracula
- Encyclia
- Epidendrum
- Epipactis
- Eria
- Eulophia
- Gongora
- Goodyera
- Grammatophyllum
- Gymnadenia
- Habenaria
- Herschelia
- Ida
- Laelia
- Lepanthes
- Liparis
- Ludisia
- Lycaste
- Renantherella
- Restrepia
- Restrepiella
- Rhynchostylis
- Roezliella
- Saccolabium
- Sarcochilus
- Satyrium
- Selenipedium
- Serapias
- Sophronitis
- Spiranthes
- Stanhopea
- Stelis
- Thrixspermum
- Tolumnia
- Trias (genus)
- Trichocentrum
- Trichoglottis
- Vanda
- Vanilla
- Yoania
- Zeuxine
- Zygopetalum
Gallery
-

Corallorhiza maculata, a myco-heterotroph
-

Cephalanthera longifolia, a terrestrial orchid
-

Leaves of different species of orchids
-

Habenaria radiata - note the lip
-

Pterostylis coccinea, a highly specialized shape
-

Neuwiedia griffithii, Apostasioideae - note the three normal stamens
-

Cypripedium acaule has two stamens. One can be seen from the picture, the other is on the other side.
-

Ophrys insectifera
-

Catasetum fimbriatum - the seta is evident.
-

Holcoglossum kimballianum
-

Vanilla plant (the climber)
-

Cattleya intermedia
-

Cultivated Epidendrum ciliare
-

Rossioglossum grande
-

Listera ovata, a less showy orchid
-

Vanda tricolor var. suavis
-

Oncidium papilio
-

Vuylstekeara- 'Cambria', an intergeneric cultivar
-

Phalaenopsis hybrid
-

Orchids at a flower show in Tatton Park, Cheshire, England, 24 July 2008
-

Renanthera storei
-

Neottia nidus-avis, a myco-heterotroph
-

An orchid leaf
-

Orchid on display at American Orchid Society, Delray Beach
-

Purple orchids at American Orchid Society, Delray Beach, FL
-

Orchid at American Orchid Society, Delray Beach
See Also
- Adaptation., a movie based on The Orchid Thief
- American Orchid Society
- Distribution of orchid species
- Italian Group for Research on Wild Orchid
- Lantingji Xu, introduction to the Orchid Pavilion Collection from Fourth Century China
- Moyobamba, known as the 'City of Orchids', which has some 3,500 species of orchid native to the area
- Nero Wolfe, a fictional detective and orchidophile
- Orchidelirium the Victorian era of flower madness in which collecting and discovering Orchid reached extraordinary levels.
- Kew Gardens video Watch the first known instance of a cricket as an orchid pollinator
- The Orchid Thief, a nonfiction book written by Susan Orlean
References
- ^ Stevens, P. F. (2001 onwards). Angiosperm Phylogeny Website Version 9, June 2008 Mobot.org
- ^ "WCSP". World Checklist of Selected Plant Families. http://apps.kew.org/wcsp/. Retrieved 2010.
- ^ Taxonomic exaggeration and its effects on orchid conservation
- ^ Corominas, Joan. "Breve Diccionario Etimológico de la Lengua Castellana". Ed. Gredos, 1980. ISBN 8424913329, pp 328
- ^ "Orchid". The Online Etymology Dictionary. http://www.etymonline.com/index.php?term=orchid. Retrieved 2010-03-25.
- ^ King, Jenny. "The coralroot orchid". Orchids in Northern Washington State. Silvercrown Mountain Outdoor School. http://www.exploresmos.org/nature/orchids-northern-washington-state/. Retrieved 10 June 2011.
- ^ a b Santiago R. Ramírez, Barbara Gravendeel, Rodrigo B. Singer, Charles R. Marshall & Naomi E. Pierce (30 August 2007). "Dating the origin of the Orchidaceae from a fossil orchid with its pollinator". Nature 448 (7157): 1042–1042. doi:10.1038/nature06039. PMID 17728756. http://www.nature.com/nature/journal/v448/n7157/abs/nature06039.html.
- ^ The origin and biogeography of Orchidaceae. In Pridgeon, AM , Cribb, PJ, Chase, MW, and Rasmussen, F, eds. Genera orchidacearum. Vol. 2. pp. 1-5 Oxford University Press , Oxford
- ^ Whitman, Melissa; Medler, Michael; Randriamanindry, Jean Jacques; Rabakonadrianina, Elisabeth (April 2011). "Conservation of Madagascar's granite outcrop orchids: influence of fire and moisture". Lankesteriana (Jardín Botánico Lankester, Universidad de Costa Rica.) 11:1: 55–67. ISSN 1409-3871. http://lankesteriana.org/lankesteriana/LANKESTERIANA%2011%281%29/9_Whitman%20et%20al.%202011.pdf.
- ^ Leake JR. 2005. Plants parasitic on fungi: unearthing the fungi in myco-heterotrophs and debunking the 'saprophytic' plant myth. Mycologist 19: 113–122. (abstract)).
- ^ Hossain MM.,"Therapeutic orchids: traditional uses and recent advances--an overview." Fitoterapia. 2011 Mar;82(2):102-40
External links
- Orchidaceae at The Plant List
- Orchidaceae at The Families of Flowering Plants (DELTA)
- Orchidaceae at the Angiosperm Phylogeny Website
- Orchidaceae at the online Flora of North America
- Orchidaceae at the online Flora of China
- Orchidaceae at the online Guide to the Flora of Mongolia
- Orchidaceae at the online Flora of Zimbabwe
- Orchidaceae at the online Flora of the Western Australian
- Orchidaceae at the online Flora of New Zealand
- Orchidaceae at Chileflora
- EPIDENDRA The Global Orchid Information Network
- Orchid Tree of Life at the Florida Museum of Natural History
- World Orchid Iconography at the Swiss Orchid Foundation
- Australian Native Orchids on Flickr
- Dr. C.E. Bracker Orchid Photographs at Ball State University.
- International Orchid Register at the Royal Horticultural Society (RHS)
- Orchid Conservation Coalition A grassroots movement towards orchid conservation
Categories:- Flowers
- Orchids
- Asparagales families
- Plants and pollinators







































Wikimedia Foundation. 2010.