- Chaperone (protein)
-
In molecular biology, chaperones are proteins that assist the non-covalent folding or unfolding and the assembly or disassembly of other macromolecular structures, but do not occur in these structures when the structures are performing their normal biological functions having completed the processes of folding and/or assembly. The common perception that chaperones are concerned primarily with protein folding is incorrect. The first protein to be called a chaperone assists the assembly of nucleosomes from folded histones and DNA and such assembly chaperones, especially in the nucleus,[1][2] are concerned with the assembly of folded subunits into oligomeric structures.[3]
Chaperones do not necessarily convey steric information required for proteins to fold: Thus, statements of the form 'chaperones fold proteins' can be misleading. One major function of chaperones is to prevent both newly synthesised polypeptide chains and assembled subunits from aggregating into nonfunctional structures. It is for this reason that many chaperones, but by no means all, are also heat shock proteins because the tendency to aggregate increases as proteins are denatured by stress. However, 'steric chaperones' directly assist in the folding of specific proteins by providing essential steric information, e.g. prodomains of bacterial proteases, lipase-specific foldases, or chaperones in fimbrial adhesion systems.
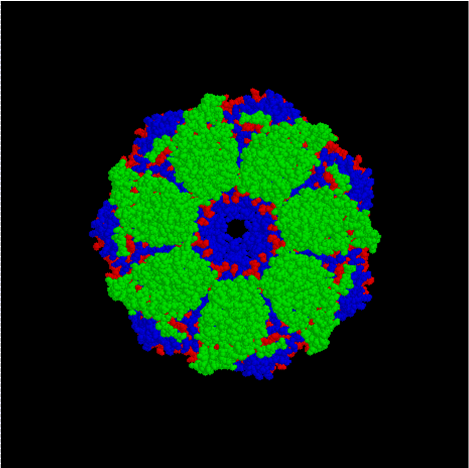
 The crystal structure of the chaperonin.
The crystal structure of the chaperonin.
Contents
Location and functions
Many chaperones are heat shock proteins, that is, proteins expressed in response to elevated temperatures or other cellular stresses.[4] The reason for this behaviour is that protein folding is severely affected by heat and, therefore, some chaperones act to repair the potential damage caused by misfolding. Other chaperones are involved in folding newly made proteins as they are extruded from the ribosome. Although most newly synthesized proteins can fold in absence of chaperones, a minority strictly requires them for the same.
Macromolecular crowding may be important in chaperone function. The crowded environment of the cytosol can accelerate the folding process, since a compact folded protein will occupy less volume than an unfolded protein chain.[5] However, crowding can reduce the yield of correctly-folded protein by increasing protein aggregation.[6][7] Crowding may also increase the effectiveness of the chaperone proteins such as GroEL,[8] which could counteract this reduction in folding efficiency.[9]
More information on the various types and mechanisms of a subset of chaperones that encapsulate their folding substrates can be found in the article for chaperonins. Chaperonins are characterized by a stacked double-ring structure and are found in prokaryotes, in the cytosol of eukaryotes, and in mitochondria.
Other types of chaperones are involved in transport across membranes, for example membranes of the mitochondria and endoplasmic reticulum (ER) in eukaryotes. Bacterial translocation—specific chaperone[10] maintains newly synthesized precursor polypeptide chains in a translocation-competent (generally unfolded) state and guides them to the translocon.
New functions for chaperones continue to be discovered, such as assistance in protein degradation, bacterial adhesin activity, and in responding to diseases linked to protein aggregation (e.g. see prion).
Human chaperone proteins
Chaperones are found in, for example, the endoplasmic reticulum (ER), since protein synthesis often occurs in this area.
Endoplasmic reticulum
In the endoplasmic reticulum (ER) there are general, lectin- and non-classical molecular chaperones helping to fold proteins.
- General chaperones: BiP, GRP94, GRP170.
- Lectin chaperones: calnexin and calreticulin
- Non-classical molecular chaperones: HSP47 and ERp29
- Folding chaperones:
- Protein disulfide isomerase (PDI),[11]
- Peptidyl prolyl cis-trans-isomerase (PPI),[12]
- ERp57[13]
Nomenclature and examples of bacterial and archeal chaperones
There are many different families of chaperones; each family acts to aid protein folding in a different way. In bacteria like E. coli, many of these proteins are highly expressed under conditions of high stress, for example, when placed in high temperatures. For this reason, the term "heat shock protein" has historically been used to name these chaperones. The prefix "Hsp" designates that the protein is a heat shock protein.
Hsp60
Hsp60 (GroEL/GroES complex in E. coli) is the best characterized large (~ 1 MDa) chaperone complex. GroEL is a double-ring 14mer with a greasy hydrophobic patch at its opening; it is so large it can accommodate native folding of 54-kDa GFP in its lumen. GroES is a single-ring heptamer that binds to GroEL in the presence of ATP or ADP. GroEL/GroES may not be able to undo previous aggregation, but it does compete in the pathway of misfolding and aggregation.[14] Also acts in mitochondrial matrix as molecular chaperone.
Hsp70
Hsp70 (DnaK in E. coli) is perhaps the best characterized small (~ 70 kDa) chaperone.
The Hsp70 proteins are aided by Hsp40 proteins (DnaJ in E. coli), which increase the ATP consumption rate and activity of the Hsp70s.
It has been noted that increased expression of Hsp70 proteins in the cell results in a decreased tendency toward apoptosis.
Although a precise mechanistic understanding has yet to be determined, it is known that Hsp70s have a high-affinity bound state to unfolded proteins when bound to ADP, and a low-affinity state when bound to ATP.
It is thought that many Hsp70s crowd around an unfolded substrate, stabilizing it and preventing aggregation until the unfolded molecule folds properly, at which time the Hsp70s lose affinity for the molecule and diffuse away.[15] Hsp70 also acts as a mitochondrial and chloroplastic molecular chaperone in eukaryotes.
Hsp90
Hsp90 (HtpG in E. coli) may be the least understood chaperone. Its molecular weight is about 90 kDa, and it is necessary for viability in eukaryotes (possibly for prokaryotes as well).
Heat shock protein 90 (Hsp90) is a molecular chaperone essential for activating many signaling proteins in the eukaryotic cell.
Each Hsp90 has an ATP-binding domain, a middle domain, and a dimerization domain. Originally thought to clamp onto their substrate protein (also known as a client protein) upon binding ATP, the recently published structures by Vaughan et al. and Ali et al. indicate that client proteins may bind externally to both the N-terminal and middle domains of Hsp90.[16][17]
Hsp90 may also require co-chaperones-like immunophilins, Sti1, p50 (Cdc37), and Aha1, and also cooperates with the Hsp70 chaperone system.[18][19]
Hsp100
Hsp100 (Clp family in E. coli) proteins have been studied in vivo and in vitro for their ability to target and unfold tagged and misfolded proteins.
Proteins in the Hsp100/Clp family form large hexameric structures with unfoldase activity in the presence of ATP. These proteins are thought to function as chaperones by processively threading client proteins through a small 20 Å (2 nm) pore, thereby giving each client protein a second chance to fold.
Some of these Hsp100 chaperones, like ClpA and ClpX, associate with the double-ringed tetradecameric serine protease ClpP; instead of catalyzing the refolding of client proteins, these complexes are responsible for the targeted destruction of tagged and misfolded proteins.
Hsp104, the Hsp100 of Saccharomyces cerevisiae, is essential for the propagation of many yeast prions. Deletion of the HSP104 gene results in cells that are unable to propagate certain prions.
History
The investigation of chaperones has a long history.[20] The term `molecular chaperone` appeared first in the literature in 1978, and was invented by Ron Laskey to describe the ability of a nuclear protein called nucleoplasmin to prevent the aggregation of folded histone proteins with DNA during the assembly of nucleosomes.[21] The term was later extended by R. John Ellis in 1987 to describe proteins that mediated the post-translational assembly of protein complexes.[22] In 1988, it was realised that similar proteins mediated this process in both prokaryotes and eukaryotes.[23] The details of this process were determined in 1989, when the ATP-dependent protein folding was demonstrated in vitro.[24]
References
- ^ Richardson RT, Alekseev OM, Grossman G et al. (July 2006). "Nuclear Autoantigenic Sperm Protein (NASP), a Linker Histone Chaperone That is Required for Cell Proliferation". Journal of Biological Chemistry 281 (30): 21526–34. doi:10.1074/jbc.M603816200. PMID 16728391.
- ^ Alekseev OM, Richardson RT, Alekseev O, O'Rand MG (2009). "Analysis of gene expression profiles in HeLa cells in response to overexpression or siRNA-mediated depletion of NASP". Reproductive Biology and Endocrinology 7: 45. doi:10.1186/1477-7827-7-45. PMC 2686705. PMID 19439102. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2686705.
- ^ Ellis RJ (July 2006). "Molecular chaperones: assisting assembly in addition to folding". Trends in Biochemical Sciences 31 (7): 395–401. doi:10.1016/j.tibs.2006.05.001. PMID 16716593.
- ^ Ellis RJ, van der Vies SM (1991). "Molecular chaperones". Annu. Rev. Biochem. 60: 321–47. doi:10.1146/annurev.bi.60.070191.001541. PMID 1679318.
- ^ van den Berg B, Wain R, Dobson CM, Ellis RJ (August 2000). "Macromolecular crowding perturbs protein refolding kinetics: implications for folding inside the cell". EMBO J. 19 (15): 3870–5. doi:10.1093/emboj/19.15.3870. PMC 306593. PMID 10921869. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=306593.
- ^ van den Berg B, Ellis RJ, Dobson CM (December 1999). "Effects of macromolecular crowding on protein folding and aggregation". EMBO J. 18 (24): 6927–33. doi:10.1093/emboj/18.24.6927. PMC 1171756. PMID 10601015. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1171756.
- ^ Ellis RJ, Minton AP (May 2006). "Protein aggregation in crowded environments". Biol. Chem. 387 (5): 485–97. doi:10.1515/BC.2006.064. PMID 16740119.
- ^ Martin J, Hartl FU (February 1997). "The effect of macromolecular crowding on chaperonin-mediated protein folding". Proc. Natl. Acad. Sci. U.S.A. 94 (4): 1107–12. doi:10.1073/pnas.94.4.1107. PMC 19752. PMID 9037014. http://www.pnas.org/cgi/pmidlookup?view=long&pmid=9037014.
- ^ Ellis RJ (2007). "Protein misassembly: macromolecular crowding and molecular chaperones". Adv. Exp. Med. Biol.. Advances in Experimental Medicine and Biology 594: 1–13. doi:10.1007/978-0-387-39975-1_1. ISBN 978-0-387-39974-4. PMID 17205670.
- ^ Zhou J, Xu Z (2005). "The structural view of bacterial translocation-specific chaperone SecB: implications for function". Molecular Microbiology 58 (2): 349–57. doi:10.1111/j.1365-2958.2005.04842.x. PMID 16194224.
- ^ Ruoppolo M, Orrù S, Talamo F, Ljung J, Pirneskoski A, Kivirikko KI, Marino G, Koivunen P (May 2003). "Mutations in domain a′ of protein disulfide isomerase affect the folding pathway of bovine pancreatic ribonuclease A". Protein Sci. 12 (5): 939–52. doi:10.1110/ps.0242803. PMC 2323865. PMID 12717017. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2323865.
- ^ Soluble complexes of target proteins and peptidyl prolyl isomerase ...
- ^ Frickel EM, Riek R, Jelesarov I, Helenius A, Wuthrich K, Ellgaard L (February 2002). "TROSY-NMR reveals interaction between ERp57 and the tip of the calreticulin P-domain". Proc. Natl. Acad. Sci. U.S.A. 99 (4): 1954–9. doi:10.1073/pnas.042699099. PMC 122301. PMID 11842220. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=122301.
- ^ Fenton WA, Horwich AL (May 2003). "Chaperonin-mediated protein folding: fate of substrate polypeptide". Q. Rev. Biophys. 36 (2): 229–56. doi:10.1017/S0033583503003883. PMID 14686103.
- ^ Mayer MP, Bukau B (March 2005). "Hsp70 chaperones: Cellular functions and molecular mechanism". Cell. Mol. Life Sci. 62 (6): 670–84. doi:10.1007/s00018-004-4464-6. PMC 2773841. PMID 15770419. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2773841.
- ^ Vaughan CK, Gohlke U, Sobott F et al. (September 2006). "Structure of an Hsp90-Cdc37-Cdk4 complex". Mol. Cell 23 (5): 697–707. doi:10.1016/j.molcel.2006.07.016. PMID 16949366. http://linkinghub.elsevier.com/retrieve/pii/S1097-2765(06)00497-7.
- ^ Ali MM, Roe SM, Vaughan CK et al. (April 2006). "Crystal structure of an Hsp90-nucleotide-p23/Sba1 closed chaperone complex". Nature 440 (7087): 1013–7. doi:10.1038/nature04716. PMID 16625188.
- ^ Terasawa K, Minami M, Minami Y (2005). "Constantly updated knowledge of Hsp90". J. Biochem. (Tokyo) 137 (4): 443–7. doi:10.1093/jb/mvi056. PMID 15858167.
- ^ Pearl LH, Prodromou C (2006). "Structure and mechanism of the Hsp90 molecular chaperone machinery". Annu. Rev. Biochem. 75: 271–94. doi:10.1146/annurev.biochem.75.103004.142738. PMID 16756493. http://arjournals.annualreviews.org/doi/full/10.1146/annurev.biochem.75.103004.142738?url_ver=Z39.88-2003&rfr_id=ori:rid:crossref.org&rfr_dat=cr_pub%3dpubmed.
- ^ Ellis RJ (1996). "Discovery of molecular chaperones". Cell Stress Chaperones 1 (3): 155–60. doi:10.1379/1466-1268(1996)001<0155:DOMC>2.3.CO;2. PMC 248474. PMID 9222600. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=248474.
- ^ Laskey RA, Honda BM, Mills AD, Finch JT (1978). "Nucleosomes are assembled by an acidic protein that binds histones and transfers them to DNA". Nature 275 (5679): 416–20. doi:10.1038/275416a0. PMID 692721.
- ^ Ellis J (1987). "Proteins as molecular chaperones". Nature 328 (6129): 378–9. doi:10.1038/328378a0. PMID 3112578.
- ^ Hemmingsen SM, Woolford C, van der Vies SM et al. (1988). "Homologous plant and bacterial proteins chaperone oligomeric protein assembly". Nature 333 (6171): 330–4. doi:10.1038/333330a0. PMID 2897629.
- ^ Goloubinoff P, Christeller JT, Gatenby AA, Lorimer GH (1989). "Reconstitution of active dimeric ribulose bisphosphate carboxylase from an unfoleded state depends on two chaperonin proteins and Mg-ATP". Nature 342 (6252): 884–9. doi:10.1038/342884a0. PMID 10532860.
See also
Chaperones/
protein foldingOtherProtein targeting Ubiquitin E1 Ubiquitin-activating enzyme (UBA1, UBA2, UBA3, UBA5, UBA6, UBA7, ATG7, NAE1, SAE1)
E2 Ubiquitin-conjugating enzyme (A • B • C • D1, D2, D3 • E1, E2, E3 • G1, G2 • H • I • J1, J2 • K • L1, L2, L3, L4, L6 • M • N • O • Q1, Q2 • R1 (CDC34), R2 • S • V1, V2 • Z)
E3 Ubiquitin ligase (VHL, Cullin, CBL, MDM2, FANCL, UBR1)
Deubiquitinating enzyme: Ataxin 3 • USP6 • CYLD
ATG3 • BIRC6 • UFC1Other Categories:- Proteins
- Protein biosynthesis

Wikimedia Foundation. 2010.