- Bipedalism
-
"Bipedality" redirects here. For the film, see Bipedality (film).
 An ostrich, one of the fastest of living bipeds
An ostrich, one of the fastest of living bipeds
 A Man Running - Eadweard Muybridge
A Man Running - Eadweard MuybridgeBipedalism is a form of terrestrial locomotion where an organism moves by means of its two rear limbs, or legs. An animal or machine that usually moves in a bipedal manner is known as a biped (
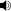 /ˈbaɪpɛd/), meaning "two feet" (from the Latin bi for "two" and ped for "foot"). Types of bipedal movement include walking, running, or hopping, on two appendages (typically legs).
/ˈbaɪpɛd/), meaning "two feet" (from the Latin bi for "two" and ped for "foot"). Types of bipedal movement include walking, running, or hopping, on two appendages (typically legs).Relatively few modern species are habitual bipeds whose normal method of locomotion is two-legged. Within mammals, habitual bipedalism has evolved six times, with the macropods, kangaroo mice, dipodids, springhare [1], pangolins and homininan apes. In the Triassic period some groups of archosaurs (a group that includes the ancestors of crocodiles) developed bipedalism; among their descendants the dinosaurs all the early forms and many later groups were habitual or exclusive bipeds; the birds descended from one group of exclusively bipedal dinosaurs.
A larger number of modern species are capable of bipedal movement for a short time in exceptional circumstances. Several non-archosaurian lizard species move bipedally when running, usually to escape from threats. Many animals rear up on their hind legs whilst fighting or copulating. A few animals commonly stand on their hind legs, in order to reach food, to keep watch, to threaten a competitor or predator, or to pose in courtship, but do not move bipedally.
There are two main types of bipedal locomotion: macropods, some smaller birds[citation needed], and heteromyid rodents move by hopping on both legs simultaneously; other groups, including apes and larger birds, walk or run by moving one leg at a time.[2]
Contents
Definition
The word is derived from the Latin words bi(s) 'two (2)' and ped- 'foot', as contrasted with quadruped 'four feet'.
Facultative and obligate bipedalism
Zoologists often label behaviors, including bipedalism, as "facultative" (i.e. optional) or "obligate" (the animal has no reasonable alternative). Even this distinction is not completely clear-cut — for example, humans normally walk and run in biped fashion, but almost all can crawl on hands and knees when necessary. There are even reports of humans who normally walk on all fours with their feet but not their knees on the ground, but these cases are a result of conditions such as Uner Tan syndrome — very rare genetic neurological disorders rather than normal behavior.[3] Even if one ignores exceptions caused by some kind of injury or illness, there are many unclear cases, including the fact that "normal" humans can crawl on hands and knees. This article therefore avoids the terms "facultative" and "obligate", and focuses on the range of styles of locomotion normally used by various groups of animals.
Movement
There are a number of states of movement commonly associated with bipedalism.
- Standing. Staying still on both legs. In most bipeds this is an active process, requiring constant adjustment of balance.
- Walking. One foot in front of another, with at least one foot on the ground at any time.
- Running. One foot in front of another, with periods where both feet are off the ground.
- Jumping/Hopping. Moving by a series of jumps with both feet moving together.
Bipedal animals
The great majority of living terrestrial vertebrates are quadrupeds. Among mammals, bipedalism is a normal method of ground locomotion in various groups of primates (e.g. lemurs, gibbons and Hominina), in the macropods (kangaroos, wallabies, etc.), and in a few groups of rodents, including kangaroo rats and kangaroo mice in the family Heteromyidae, as well as gerbils and spring hares). All birds are bipeds when on the ground, a feature inherited from their dinosaur ancestors. Bipedalism evolved more than once in archosaurs, the group that includes both dinosaurs and crocodilians.[4] Many species of lizards become bipedal during high-speed, sprint locomotion, including the world's fastest lizard, the spiny-tailed iguana (genus Ctenosaura). There are no known living or fossil bipedal amphibians.
Most bipedal animals move with their backs close to horizontal, using a long tail to balance the weight of their bodies. The primate version of bipedalism is unusual because the back is close to upright (completely upright in humans) and, among primates that move bipedally, only the lemurs have tails.
Humans and large birds walk by raising one foot at a time. On the other hand most macropods, smaller birds and bipedal rodents move by hopping on both legs simultaneously. Tree kangaroos are able to utilize either form of locomotion, most commonly alternating feet when moving arboreally and hopping on both feet simultaneously when on the ground.
Archosaurs (include birds, crocodiles, and dinosaurs)
Birds are bipedal when not flying, with small birds tending to hop and larger birds walking with an alternating gait. All dinosaurs are believed to be descended from a fully bipedal ancestor, perhaps similar to Eoraptor. Bipedal movement also re-evolved in a number of other dinosaur lineages such as the iguanodons. Some extinct members of the crocodilian line, a sister group to the dinosaurs and birds, also evolved bipedal forms - a crocodile relative from the triassic, Effigia okeeffeae, is believed to be bipedal.[5] Pterosaurs were previously thought to have been bipedal, but recent trackways have all shown quadrupedal locomotion.
Mammals
Bipedal movement is less common among mammals, most of which are quadrupedal. All primates possess some bipedal ability, though non-human primates primarily use quadrupedal locomotion on land. Primates aside, the largest mammalian group using exclusive bipedal movement are the macropods (kangaroos, wallabies and their relatives), which move via hopping. Other mammals also move via hopping, such as the kangaroo rat, springhare and certain primates such as the sifaka and sportive lemur. Very few mammals other than primates commonly move bipedally by an alternating gait rather than hopping. Exceptions are the ground pangolin and in some circumstances the tree kangaroo. [1]
Primates
Primates can be distinguished from other quadrupedal mammals as they exhibit a greater diversity in locomotor behaviors.[6] These include arm swinging (brachiation), quadrumanous climbing, knuckle walking, and regular short bouts of bipedalism. In addition, quadrupedal locomotion in primates also exhibits significant differences from other mammals. These differences in gait characteristics are primarily adaptations to an arboreal environment.[7] All primates can sit upright. Many primates can stand upright on their hind legs without any support. Chimpanzees, bonobos, gibbons[8] and baboons[9] exhibit relatively advanced forms of bipedalism. Injured chimpanzees and bonobos have been capable of sustained bipedalism.[10]
Primates that live in tropical areas often wade through water in a bipedal stance. Gorillas, bonobos, proboscis monkeys and baboons have been observed wading bipedally. Three captive primates, one macaque Natasha[11] and two chimps, Oliver and Poko (chimpanzee), were found to move bipedally. Natasha switched to exclusive bipedalism after an illness, while Poko was discovered in captivity in a tall, narrow cage.[12][13] Oliver reverted to knuckle-walking after developing arthritis.
In addition, non-human primates often use bipedal locomotion when carrying food. One hypothesis for human bipedalism is thus that it evolved as a result of differentially successful survival from carrying food to share with group members,[14] although there are other hypotheses, as discussed below.
Limited bipedalism in mammals
Other mammals engage in limited, non-locomotory, bipedalism. A number of other animals, such as rats, racoons, and beavers will squat on their hindlegs to manipulate some objects but revert to four limbs when moving (the beaver may also move bipedally if transporting wood for their dams). Bears will fight in a bipedal stance to use their forelegs as weapons. Ground squirrels and meerkats will stand on hind legs to survey their surroundings, but will not walk bipedally. Dogs can stand or move on two legs if trained, or if birth defect or injury precludes quadrupedalism. The gerenuk antelope stands on its hind legs while eating from trees, as did the extinct giant ground sloth and chalicotheres. The spotted skunk will also use limited bipedalism when threatened, rearing up on its forelimbs while facing the attacker so its anal glands, capable of spraying an offensive oil, face its attacker.
Limited bipedalism in non-mammals
Bipedalism is unknown among the amphibians. Among the non-archosaur reptiles bipedalism is rare, but it is found in the 'reared-up' running of lizards such as agamids and monitor lizards. Many reptile species will also temporarily adopt bipedalism while fighting.[15] One genus of basilisk lizard can run bipedally across the surface of water for some distance. Among arthropods, cockroaches are known to move bipedally at high speeds [2]. Bipedalism is virtually solely found in terrestrial animals, though at least two types of octopus walk bipedally on the sea floor using two of their arms, allowing the remaining arms to be used to camouflage the octopus as a mat of algae or a floating coconut.[16]
Advantages
Limited and exclusive bipedalism can offer a species several advantages. Bipedalism raises the head; this allows a greater field of vision with improved detection of distant dangers or resources, access to deeper water for wading animals and allows the animals to reach higher food sources with their mouths. While upright, non-locomotory limbs become free for other uses, including manipulation (in primates and rodents), flight (in birds), digging (in giant pangolin), combat (in bears and the large monitor lizard) or camouflage (in certain species of octopus). Running speeds can be increased when an animal lacks a flexible backbone, though the maximum bipedal speed appears less fast than the maximum speed of quadrupedal movement with a flexible backbone – the ostrich reaches speeds of 65 km/h (40 mph) and the red kangaroo 70 km/h (43 mph), while the cheetah can exceed 100 km/h (62 mph).[17] [18] Bipedality in kangaroo rats has been hypothesized to improve locomotor performance, which could aid in escaping from predators.[19][20]
Evolution
Recent evidence regarding modern human sexual dimorphism (physical differences between men and women) in the lumbar spine has been seen in pre-modern primates such as Australopithecus africanus. This dimorphism has been seen as an evolutionary adaptation of females to bear lumbar load better during pregnancy, an adaptation that non-bipedal primates would not need to make.[21][22]
Bipedalism has a number of adaptive advantages, and has evolved independently in a number of lineages.
Early reptiles and lizards
The first known biped is the bolosaurid Eudibamus whose fossils date from 290 million years ago.[23][24] Its long hindlegs, short forelegs, and distinctive joints all suggest bipedalism. This may have given increased speed. The species was extinct before the dinosaurs appeared.
Independent of Eudibamus, some modern lizard species have developed the capacity to run on their hind legs for added speed.
Dinosaurs and birds
Bipedalism also evolved independently among the dinosaurs. Dinosaurs diverged from their archosaur ancestors approximately 230 million years ago during the Middle to Late Triassic period, roughly 20 million years after the Permian-Triassic extinction event wiped out an estimated 95% of all [life on Earth].[25][26] Radiometric dating of fossils from the early dinosaur genus Eoraptor establishes its presence in the fossil record at this time. Paleontologists believe Eoraptor resembles the common ancestor of all dinosaurs;[27] if this is true, its traits suggest that the first dinosaurs were small, bipedal predators.[28] The discovery of primitive, dinosaur-like ornithodirans such as Marasuchus and Lagerpeton in Argentinian Middle Triassic strata supports this view; analysis of recovered fossils suggests that these animals were indeed small, bipedal predators.
Mammals
A number of mammals will adopt a bipedal stance in specific situations such as for feeding or fighting. A number of groups of extant mammals have independently evolved bipedalism as their main form of locomotion - for example humans, giant pangolins, the extinct giant ground sloths, numerous species of jumping rodents and macropods. Humans, as their bipedalism has been extensively studied are documented in the next section. Macropods are believed to have evolved bipedal hopping only once in their evolution, at some time no later than 45 million years ago.[29]
Humans
Main article: Human skeletal changes due to bipedalismThere are at least twelve distinct hypotheses as to how and why bipedalism evolved in humans, and also some debate as to when. Bipedalism evolved well before the large human brain or the development of stone tools.[30] Bipedal specializations are found in Australopithecus fossils from 4.2-3.9 million years ago [31] The different hypotheses are not necessarily mutually exclusive and a number of selective forces may have acted together to lead to human bipedalism. It is important to distinguish between adaptations for bipedalism and adaptations for running, which came later still.
Possible reasons for the evolution of human bipedalism include freeing the hands for tool use and carrying, sexual dimorphism in food gathering, changes in climate and habitat (from jungle to savanna) and to reduce the amount of skin exposed to the tropical sun.
Traveling efficiency hypothesis
An alternative explanation is the mixture of savanna and scattered forests increased terrestrial travel by proto-humans between clusters of trees, and bipedalism offered greater efficiency for long-distance travel between these clusters than quadrupedalism.[32][33]
Postural feeding hypothesis
The postural feeding hypothesis has been recently supported by Dr. Kevin Hunt, a professor at Indiana University. This hypothesis asserts that chimpanzees were only bipedal when they ate. While on the ground, they would reach up for fruit hanging from small trees and while in trees, bipedalism was utilized by grabbing for an overhead branch. These bipedal movements may have evolved into regular habits because they were so convenient in obtaining food. Also, Hunt hypothesises that these movements coevolved with chimpanzee arm-hanging, as this movement was very effective and efficient in harvesting food. When analyzing fossil anatomy, Australopithecus afarensis has very similar features of the hand and shoulder to the chimpanzee, which indicates hanging arms. Also, the Australopithecus hip and hind limb very clearly indicate bipedalism, but these fossils also indicate very inefficient locomotive movement when compared to humans. For this reason, Hunt argues that bipedalism evolved more as a terrestrial feeding posture than as a walking posture. As Hunt says, “A bipedal postural feeding adaptation may have been a preadaptation for the fully realized locomotor bipedalism apparent in Homo erectus.”[citation needed]
Provisioning model
One theory on the origin of bipedalism is the behavioral model presented by C. Owen Lovejoy, known as "male provisioning".[34] Lovejoy theorizes that the evolution of bipedalism was linked to monogamy. In the face of long inter-birth intervals and low reproductive rates typical of the apes, early hominids engaged in pair-bonding that enabled greater parental effort directed towards rearing offspring. Lovejoy proposes that male provisioning of food would improve the offspring survivorship and increase the pair's reproductive rate. Thus the male would leave his mate and offspring to search for food and return carrying the food in his arms walking on his legs. This model is supported by the reduction ("feminization") of the male canine teeth in early hominids such as Sahelanthropus tchadensis [35] and Ardipithecus ramidus,[36] which along with low body size dimorphism in Ardipithecus[37] and Australopithecus,[38] suggests a reduction in inter-male antagonistic behavior in early hominids.[39] In addition, this model is supported by a number of modern human traits associated with concealed ovulation (permanently enlarged breasts, lack of sexual swelling) and low sperm competition (moderate sized testes, low sperm mid-piece volume) that argues against recent adaptation to a polygynous reproductive system.[39]
However, this model has generated some controversy, as others have argued that early bipedal hominids were instead polygynous. Among most monogamous primates, males and females are about the same size. That is sexual dimorphism is minimal, and other studies have suggested that Australopithecus afarensis males were nearly twice the weight of females (as well as a great deal taller)[citation needed]. However, Lovejoy's model posits that the larger range a provisioning male would have to cover (to avoid competing with the female for resources she could attain herself) would select for increased male body size to limit predation risk.[40] Furthermore, as the species became more bipedal, specialized feet would prevent the infant from conveniently clinging to the mother - hampering the mother's freedom [41] and thus make her and her offspring more dependent on resources collected by others. Modern monogamous primates such as gibbons tend to be also territorial, but fossil evidence indicates that Australopithecus afarensis lived in large groups. However, while both gibbons and hominids have reduced canine sexual dimorphism, female gibbons enlarge ('masculinize') their canines so they can actively share in the defense of their home territory. Instead, the reduction of the male hominid canine is consistent with reduced inter-male aggression in a group living primate.
Early bipedalism in homininae model
 Evolution of bipedalism in homininae
Evolution of bipedalism in homininaeRecent studies of 4.4 million years old Ardipithecus ramidus suggest bipedalism, it is thus possible that bipedalism evolved very early in homininae and was reduced in chimpanzee and gorilla when they became more specialized. According to Richard Dawkins in his book "The Ancestor's Tale", chimps and bonobos are descended from Australopithecus gracile type species while gorillas are descended from Paranthropus. These apes may have once been bipedal, but then lost this ability when they were forced back into an arboreal habitat, presumably by those australopithecines who eventually became us (see Homininae). Early homininaes such as Ardipithecus ramidus may have possessed an arboreal type of bipedalism that later independently evolved towards knuckle-walking in chimpanzees and gorillas [42] and towards efficient walking and running in modern humans (see figure). It is also proposed that one cause of Neanderthal extinction was a less efficient running.
Warning display (Aposematic) model
Joseph Jordania from the University of Melbourne recently (2011) suggested that bipedalism was one of the central element of the general defense strategy of early hominids, based on aposematism, or warning display and intimidation of potential predators and competitors with exaggerated visual and audio signals. According to this model, hominids were trying to stay as visible and as loud as possible all the time. Several morphological and behavioral developments were employed to achieve this goal: upright bipedal posture, longer legs, long tightly coiled hair on the top of the head, body painting, threatening synchronous body movements, loud voice and extremely loud rhythmic singing/stomping/drumming on external subjects [43]. Slow locomotion and strong body odor (both characteristic for hominids and humans) are other features often employed by aposematic species to advertise their non-profitability for potential predators.
Other behavioural models
There are a variety of ideas which promote a specific change in behaviour as the key driver for the evolution of hominid bipedalism. For example, Wescott (1967) and later Jablonski & Chaplin (1993) suggest that bipedal threat displays could have been the transitional behaviour which led to some groups of apes beginning to adopt bipedal postures more often. Others (e.g. Dart 1925) have offered the idea that the need for more vigilance against predators could have provided the initial motivation. Dawkins (e.g. 2004) has argued that it could have begun as a kind of fashion that just caught on and then escalated through sexual selection. And it has even been suggested (e.g. Tanner 1981:165) that male phallic display could have been the initial incentive.
Thermoregulatory model
The thermoregulatory model explaining the origin of bipedalism is one of the simplest theories so far advanced, but it is a viable explanation. Dr. Peter Wheeler, a professor of evolutionary biology, proposes that bipedalism raises the amount of body surface area higher above the ground which results in a reduction in heat gain and helps heat dissipation. When a hominid is higher above the ground, the organism accesses more favorable wind speeds and temperatures. During heat seasons, greater wind flow results in a higher heat loss, which makes the organism more comfortable. Also, Wheeler explains that a vertical posture minimizes the direct exposure to the sun whereas quadrupedalism exposes more of the body to direct exposure. Analysis and interpretations of Ardipithecus reveal that this hypothesis needs modification to consider that the forest and woodland environmental preadaptation of early-stage hominid bipedalism preceded further refinement of bipedalism by the pressure of natural selection. This then allowed for the more efficient exploitation of the hotter conditions ecological niche, rather than the hotter conditions being hypothetically bipedalism's initial stimulus.
Carrying models
Charles Darwin wrote that "Man could not have attained his present dominant position in the world without the use of his hands, which are so admirably adapted to the act of obedience of his will" Darwin (1871:52) and many models on bipedal origins are based on this line of thought. Gordon Hewes (1961) suggested that the carrying of meat "over considerable distances" (Hewes 1961:689) was the key factor. Isaac (1978) and Sinclair et al. (1986) offered modifications of this idea as indeed did Lovejoy (1981) with his 'provisioning model' described above. Others, such as Nancy Tanner (1981) have suggested that infant carrying was key, whilst others have suggested stone tools and weapons drove the change.[who?]
Wading models
Several theories have been proposed regarding the influence of water on human bipedalism. The aquatic ape hypothesis, promoted for several decades by Elaine Morgan, proposed that swimming, diving and aquatic food sources exerted a strong influence on many aspects of human evolution, including bipedalism.[44] It is not accepted by or considered a serious theory within anthropological scholarly community.[45][46]
Other scholarly theories have been proposed that suggest wading[47] and the exploitation of aquatic food sources (providing essential nutrients for human brain evolution[48] or critical fallback foods[49]) may have exerted evolutionary pressures on human ancestors leading to bipedalism.
Physiology
Bipedal movement occurs in a number of ways, and requires many mechanical and neurological adaptations. Some of these are described below.
Biomechanics
Standing
Energy-efficient means of standing bipedally involve constant adjustment of balance, and of course these must avoid overcorrection. The difficulties associated with simple standing in upright humans are highlighted by the greatly increased risk of falling present in the elderly, even with minimal reductions in control system effectiveness.
Walking
Walking is characterized by an "inverted pendulum" movement in which the body vaults over a stiff leg with each step.[50] Force plates can be used to quantify the whole-body kinetic & potential energy, with walking displaying an out-of-phase relationship indicating exchange between the two.[50] Interestingly, this model applies to all walking organisms regardless of the number of legs, and thus bipedal locomotion does not differ in terms of whole-body kinetics.[51]
In humans, walking is composed of several separate processes[50]:
- Vaulting over a stiff stance leg
- Passive ballistic movement of the swing leg
- A short 'push' from the ankle prior to toe-off, propelling the swing leg
- Rotation of the hips about the axis of the spine, to increase stride length
- Rotation of the hips about the horizontal axis to improve balance during stance
Running
Running is characterized by a spring-mass movement.[50] Kinetic and potential energy are in phase, and the energy is stored & released from a spring-like limb during foot contact.[50] Again, the whole-body kinetics are similar to animals with more limbs.[51]
Musculature
Bipedalism requires strong leg muscles, particularly in the thighs. Contrast in domesticated poultry the well muscled legs, against the small and bony wings. Likewise in humans, the quadriceps and hamstring muscles of the thigh are both so crucial to bipedal activities that each alone is much larger than even the well-developed biceps of the arms.
Nervous system
The famous knee jerk (or patellar reflex) emphasizes the necessary bipedal control system: the only function served by the nerves involved being connected as they are is to ensure quick response to imminent disturbance of erect posture; it not only occurs without conscious mental activity, but also involves none of the nerves which lead from the leg to the brain.[citation needed]
A less well-known aspect of bipedal neuroanatomy can be demonstrated in human infants who have not yet developed toward the ability to stand up. They can nevertheless run with great dexterity, provided they are supported in a vertical position and offered the stimulus of a moving treadmill beneath their feet.[citation needed]
Respiration
A biped also has the ability to breathe whilst it runs. Humans usually take a breath every other stride when their aerobic system is functioning. During a sprint, at which point the anaerobic system kicks in, breathing slows until the anaerobic system can no longer sustain a sprint.
This is not necessarily an advantage over quadrupeds, as not only can many quadrupeds breathe while running, but in mammals such as dogs, the act of running helps to expand and contract the lungs. The muscles of the trunk thus perform locomotive and respiratory tasks at the same time, making breathing while running more efficient in these animals than in bipeds.[citation needed].
Bipedal robots
Main article: humanoid robot ASIMO - a bipedal robot
ASIMO - a bipedal robotFor nearly the whole of the 20th century, bipedal robots were very difficult to construct and robot locomotion involved only wheels, treads, or multiple legs. Recent cheap and compact computing power has made two-legged robots more feasible. Some notable biped robots are ASIMO, HUBO and QRIO. Recently, spurred by the success of creating a fully passive, un-powered bipedal walking robot,[52] those working on such machines have begun using principles gleaned from the study of human and animal locomotion, which often relies on passive mechanisms to minimize power consumption.
See also
- Allometry
- Quadrupedalism
- Orthograde posture
Notes
- ^ NC Heglund, GA Cavagna and CR Taylor 1982 Energetics and mechanics of terrestrial locomotion. III. Energy changes of the centre of mass as a function of speed and body size in birds and mammals Journal of Experimental Biology 97:1
- ^ Dhingra, Philip (2004-05-25). "Comparative bipedalism: How the rest of the animal kingdom walks on two legs". http://www.philosophistry.com/static/bipedalism.html. Retrieved 2007-10-29.
- ^ Humphrey, N., Skoyles, J.R., and Keynes, R. (2005). "Human Hand-Walkers: Five Siblings Who Never Stood Up" (PDF). Centre for Philosophy of Natural and Social Science, London School of Economics. http://www.lse.ac.uk/collections/CPNSS/pdf/DP_withCoverPages/DP77/DP7705.pdf.
- ^ Hutchinson, J.R. (2006). "The evolution of locomotion in archosaurs". Comptes Rendus Palevol 5 (3–4): 519–530. doi:10.1016/j.crpv.2005.09.002. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6X1G-4J6WP8M-1&_user=10&_rdoc=1&_fmt=&_orig=search&_sort=d&view=c&_acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=ff4c612aef14e51e872112989be76277.
- ^ Handwerk, Brian (2006-01-26). "Dino-Era Fossil Reveals Two-Footed Croc Relative". National Geographic. http://news.nationalgeographic.com/news/2006/01/0125_060125_crocodile.html. Retrieved 2007-10-29.
- ^ Fleagle, J.G (1994). "Primate locomotion and posture". In Steve Jones, Robert Martin & David Pilbeam. The Cambridge Encyclopedia of Human Evolution. Cambridge: Cambridge University Press. pp. 75–79. ISBN 0-521-3270-3. Also ISBN-0-521-46786-1 (paperback)
- ^ Schmitt, Daniel (2003). "Insights into the evolution of human bipedalism from experimental studies of humans and other primates". Journal of Experimental Biology 206 (Pt 9): 1437–48. doi:10.1242/jeb.00279. PMID 12654883. http://jeb.biologists.org/cgi/content/full/206/9/1437.
- ^ Aerts, Peter; Evie E. Vereeckea, Kristiaan D'Aoûta (2006). "Locomotor versatility in the white-handed gibbon (Hylobates lar): A spatiotemporal analysis of the bipedal, tripedal, and quadrupedal gaits". Journal of Human Evolution (Elsevier) 50 (5): 552–567. doi:10.1016/j.jhevol.2005.12.011. PMID 16516949. http://www.sciencedirect.com/science?_ob=ArticleURL&_udi=B6WJS-4JD1114-1&_user=10&_rdoc=1&_fmt=&_orig=search&_sort=d&_docanchor=&view=c&_searchStrId=1152159996&_rerunOrigin=scholar.google&_acct=C000050221&_version=1&_urlVersion=0&_userid=10&md5=ee529d84e58c8ff5f5c98b5e4945dc76.
- ^ Rose, M.D. (1976). "Bipedal behavior of olive baboons (Papio anubis) and its relevance to an understanding of the evolution of human bipedalism". American Journal of Physical Anthropology 44 (2): 247–261. doi:10.1002/ajpa.1330440207. PMID 816205. http://www3.interscience.wiley.com/journal/110488308/abstract.
- ^ Bauer, Harold (1976). "Chimpanzee bipedal locomotion in the Gombe National Park, East Africa". Primates 18 (4): 913. doi:10.1007/BF02382940. http://www.springerlink.com/content/067867j341t13662/.
- ^ Waldman, Dan (2004-07-21). "Monkey apes humans by walking on two legs". MSNBC. http://www.msnbc.msn.com/id/5479501. Retrieved 2007-10-29.
- ^ University of Liverpool - Research Intelligence Issue 22 - Walking tall after all
- ^ Tetrapod Zoology : Bipedal orangs, gait of a dinosaur, and new-look Ichthyostega: exciting times in functional anatomy part I
- ^ Bipedality in chimpanzee (Pan troglodytes) and bonobo (Pan paniscus): Testing hypotheses on the evolution of bipedalism
- ^ Sharma, Jayanth (2007-03-08). "The Story behind the Picture - Monitor Lizards Combat" (php). Wildlife Times. http://www.wildlifetimes.com/forums/viewtopic.php?p=601. Retrieved 2007-10-29.
- ^ Huffard CL, Boneka F, Full RJ (2005). "Underwater bipedal locomotion by octopuses in disguise". Science 307 (5717): 1927. doi:10.1126/science.1109616. PMID 15790846.
- ^ Garland, T., Jr. (1983). "The relation between maximal running speed and body mass in terrestrial mammals". Journal of Zoology, London 199 (2): 157–170. doi:10.1111/j.1469-7998.1983.tb02087.x. http://www.biology.ucr.edu/people/faculty/Garland/Garl1983_JZL.pdf.
- ^ Sharp, N.C.C. (1997). "Timed running speed of a cheetah (Acinonyx jubatus)". Of Zoology, London 241 (3): 493–494. doi:10.1111/j.1469-7998.1997.tb04840.x.
- ^ Djawdan, M (1993). "Locomotor performance of bipedal and quadrupedal heteromyid rodents". Functional Ecology (British Ecological Society) 7 (2): 195–202. doi:10.2307/2389887. JSTOR 2389887.
- ^ Djawdan, M.; Garland, T., Jr. (1988). "Maximal running speeds of bipedal and quadrupedal rodents". Journal of Mammalogy (American Society of Mammalogists) 69 (4): 765–772. doi:10.2307/1381631. JSTOR 1381631. http://www.biology.ucr.edu/people/faculty/Garland/DjawGa88.pdf.
- ^ The Independent's article A pregnant woman's spine is her flexible friend, by Steve Connor from The Independent (Published: 13 December 2007) quoting Shapiro, Liza, University of Texas at Austin Dept. of Anthropology about her article, Whitcome, et al., Nature advance online publication, doi:10.1038/nature06342 (2007).
- ^ Why Pregnant Women Don't Tip Over. Amitabh Avasthi for National Geographic News, December 12, 2007. This article has good pictures explaining the differences between bipedal and non-bipedal pregnancy loads.
- ^ "Upright lizard leaves dinosaur standing". cnn.com. 2000-11-03. Archived from the original on 2007-10-31. http://web.archive.org/web/20071031005956/http://archives.cnn.com/2000/NATURE/11/03/germany.dinosaur/index.html. Retrieved 2007-10-17.
- ^ Berman, David S. et al. (2000). "Early Permian Bipedal Reptile". Science 290 (5493): 969–972. doi:10.1126/science.290.5493.969. PMID 11062126. http://www.sciencemag.org/cgi/content/abstract/290/5493/969.
- ^ Citation for Permian/Triassic extinction event, percentage of animal species that went extinct. See commentary
- ^ Another citation for P/T event data. See commentary
- ^ Hayward, T. (1997). The First Dinosaurs. Dinosaur Cards. Orbis Publishing Ltd. D36040612.
- ^ Sereno, Paul C.; Catherine A. Forster, Raymond R. Rogers and Alfredo M. Monetta (January 1993). "Primitive dinosaur skeleton from Argentina and the early evolution of Dinosauria". Nature 361 (6407): 64–66. doi:10.1038/361064a0. http://www.nature.com/nature/journal/v361/n6407/abs/361064a0.html. Retrieved 2008-06-28.
- ^ Burk, Angela; Michael Westerman and Mark Springer (September 1988). "The Phylogenetic Position of the Musky Rat-Kangaroo and the Evolution of Bipedal Hopping in Kangaroos (Macropodidae: Diprotodontia)". Systematic Biology 47 (3): 457–474. doi:10.1080/106351598260824. PMID 12066687.
- ^ Lovejoy, C.O. (1988). "Evolution of Human walking". Scientific American. 259 (5): 82–89.
- ^ McHenry, H.M (2009). "Human Evolution". In Michael Ruse & Joseph Travis. Evolution: The First Four Billion Years. Cambridge, Massachusetts: The Belknap Press of Harvard University Press. p. 263. ISBN 978-0-674-03175-3.
- ^ Isbell, L.A. and T.P. Young. (1996). "The evolution of bipedalism in hominids and reduced group size in chimpanzees: alternative responses to decreasing resource availability". Journal of Human Evolution 30: 289–297.
- ^ Lewin, Roger; Swisher, Carl Celso; Curtis, Garniss H. (2000). Java man: how two geologists' dramatic discoveries changed our understanding of the evolutionary path to modern humans. New York: Scribner. ISBN 0-684-80000-4.
- ^ T. Douglas Price, Gary M. Feinman (2003). Images of the Past, 5th edition. Boston: McGraw Hill. pp. 68. ISBN 978-0-07-340520-9.
- ^ Brunet, Michel; Guy F, Pilbeam D, Mackaye HT, Likius A et al. (11). "A new hominid from the Upper Miocene of Chad, Central Africa". Nature 418 (6894): 145–151. doi:10.1038/nature00879. PMID 12110880. http://www.nature.com/nature.
- ^ Suwa, Gen; Kono RT, Simpson SW, Asfaw B, Lovejoy CO, White TD (2). "Paleobiological implications of the Ardipithecus ramidus dentition". Science 326 (5949): 94–99. doi:10.1126/science.1175824. PMID 19810195. http://www.sciencemag.org.
- ^ White TD et al. Science. 2009 326(5949):75-86
- ^ Reno PL et al. Philos Trans R Soc Lond B Biol Sci. 2010 365(1556):3355-63; Harmon E. J Hum Evol. 2009 56(6):551-9.
- ^ a b Lovejoy CO. Science. 2009 326(5949):74e1-8.
- ^ Lovejoy CO. Science. 1981 211(4480):341-50.
- ^ Keith Oatley, Dacher Keltner, Jennifer M. Jenkins. Understanding Emotion (2006) Second Edition. Page 235.
- ^ Kivell TL, Schmitt D 2009 Independent evolution of knuckle-walking in African apes shows that humans did not evolve from a knuckle-walking ancestor. Proc Natl Acad Sci U S A. 2009 Aug 25;106(34):14241-6. PMID: 19667206
- ^ Joseph Jordania. Why do People Sing? Music in Human Evolution. Logos, 2011
- ^ Morgan, Elaine (1997). The Aquatic Ape Hypothesis. Souvenir Press. ISBN 0285635182.
- ^ Bridgeman, B (2003). Psychology & evolution: the origins of mind. SAGE Publications. pp. 64. ISBN 0761924795.
- ^ Meier, R (2003). The complete idiot's guide to human prehistory. Alpha Books. pp. 57–59. ISBN 0028644212.
- ^ . ISBN 0011400906373
- ^ Cunnane, Stephen C (2005). Survival of the fattest: the key to human brain evolution. World Scientific Publishing Company. pp. 259. ISBN 9812561919.
- ^ Wrangham R, Cheney D, Seyfarth R, Sarmiento E (December 2009). "Shallow-water habitats as sources of fallback foods for hominins". Am. J. Phys. Anthropol. 140 (4): 630–42. doi:10.1002/ajpa.21122. PMID 19890871.
- ^ a b c d e McMahon, Thomas A (1984). Muscles, reflexes, and locomotion. ISBN 9780691023762. http://books.google.com/?id=1sz5501URh0C&printsec=frontcover&dq=macmahon+muscles#v=onepage&q&f=false.
- ^ a b Biewener, Andrew A; Daniel, T (2003). "A moving topic: control and dynamics of animal locomotion". Biology letters 6 (3): 387–8. doi:10.1098/rsbl.2010.0294. ISBN 9780198500223. PMC 2880073. PMID 20410030. http://books.google.com/books?id=yMaN9pk8QJAC&printsec=frontcover#v=onepage&q&f=false.
- ^ http://ruina.tam.cornell.edu/hplab/pdw.html
References
- Darwin, C., "The Descent of Man and Selection in Relation to Sex", Murray (London), (1871).
- Dart, R.A., "Australopithecus africanus: The Ape Man of South Africa" Nature, 145, 195-199, (1925).
- Dawkins, R., "The Ancestor's Tale", Weidenfeld and Nicolson (London), (2004).
- Hewes, G.W., "Food Transport and the Origin of Hominid Bipedalism" American Anthropologist, 63, 687-710, (1961).
- Hunt, K.D., "The Evolution of Human Bipedality" Journal of Human Evolution, 26, 183-202, (1994).
- Isaac, G.I., "The Archeological Evidence for the Activities of Early African Hominids" In:Early Hominids of Africa (Jolly, C.J. (Ed.)), Duckworth (London), 219-254, (1978).
- Jablonski, N.G. & Chaplin, G. "Origin of Habitual Terrestrial Bipedalism in the Ancestor of the Hominidae", Journal of Human Evolution, 24, 259-280, (1993).
- Lovejoy, C. O., "The Origin of Man." Science, 211, 341-350, (1981).
- Tanner, N.M., "On Becoming Human", Cambridge University Press (Cambridge), (1981).
- Wescott, R.W., "Hominid Uprightness and Primate Display", American Anthropologist, 69, 738,(1967).
- Wheeler, P. E., "The Evolution of Bipedality and Loss of Functional Body Hair in Hominoids." Journal of Human Evolution, 13, 91-98, (1984).
- Vrba, E., "The Pulse that Produced Us." Natural History, 102(5), 47-51, (1993).
External links
Animal locomotion on land Gait class LeggedBrachiation · Arboreal locomotion · Hand-walking · Jumping · Knuckle-walking · Gait · Running · WalkingLeglessConcertina movement · Undulatory locomotion · Rectilinear locomotion · Rolling · Sidewinding · Other modesAnatomy Comparative foot morphology · Digitigrade · Plantigrade · Ungulate · Uniped · Biped · Triped · Quadruped · Facultative bipedSpecific Categories:- Locomotion
- Animal anatomy



Wikimedia Foundation. 2010.